【Nature综述】朱健康院士等综述植物非生物胁迫信号转导,为培育抗逆作物提供思路(全文翻译,值得收...
来源: 时间:2024年11月26日 11:09

Nature Reviews Genetics (IF=53.24)杂志在线发表了来自中国科学院分子植物科学卓越创新中心/上海植物逆境生物学研究中心朱健康课题组及张惠明课题组研究员共同通讯题为“Abiotic stress responses in plants”的综述文章,该综述总结了最新关于植物对非生物胁迫响应的分子机制研究进展,表明植物对非生物胁迫响应是多层次和多个过程,其中包括信号感应及传导、转录及转录调控、翻译和翻译后蛋白质修饰等。最后文章还展望了未来还可能利用这些机制通过遗传、化学和微生物方法提高作物生产力和农业可持续性。

值得注意的是,该课题组还在Cell杂志发表题为““Abiotic stress signaling and responeses in plants”的综述文章。并且已经入选高被引用文章,截止202109,其文章的被引次数为1451次。详细请参考我们公众号iPlants报道的全文翻译:Cell综述|朱健康院士综述植物非生物胁迫信号转导(全文翻译,值得收藏)。该课题组近几年依然连续在两大权威顶尖杂志上发表关于植物非生物胁迫的分子机制的综述文章,说明了该课题在植物非生物胁迫机制研究领域一直处在世界领先水平。

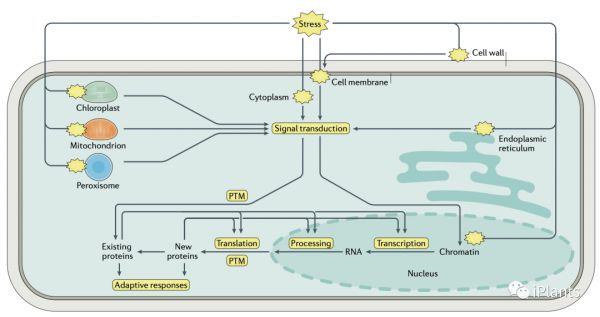
1.感应胁迫
环境胁迫可直接引起植物细胞内生物分子的物理或化学变化,从而引发细胞应激反应。但是很难证明生物分子能直接感知压力,因此大多数假定的压力传感器都是使用间接方法识别的。例如,破坏感受器的功能预计会影响第二信使的水平,例如 Ca 2+、ROS、一氧化氮和磷脂,并且已经设计了遗传筛选来识别导致此类表型的突变。特别是细胞内游离 Ca 2+浓度快速显示响应外部压力的刺激特异性模式,这使得能够在表达基于水母发光蛋白的 Ca2+指标的转基因植物的遗传筛选中研究渗透压、盐和温度压力的感知。感知渗透压的变化
干旱和盐度都会在植物细胞中诱导高渗胁迫。在拟南芥中,编码高渗门控钙通道 OSCA1 的基因在基于钙成像的遗传筛选中被鉴定为高渗压感受器(见下图 )。OSCA1 位于质膜中,其功能障碍导致保卫细胞和根细胞中Ca 2+流入减少、叶片蒸腾作用缺失和高渗胁迫下植物的根生长减少。水稻(水稻)和拟南芥中 OSCA1 家族蛋白的结构分析为它们如何充当感受器提出一种可能的机制;降低的高渗条件下细胞膨压降低在脂双层的横向张力,导致OSCA离子通道打开和Ca 2+转运入细胞。低渗透压也会改变细胞膜张力。在拟南芥中,MSL8 是花粉中低渗应激诱导的膜张力的感受器。低渗透压下增加的膜张力增加了 MSL8 打开以允许离子流出的可能性,从而保护细胞免受内部渗透压的影响。感知盐浓度的变化
除了高渗胁迫之外,在盐渍土壤中生长的植物还遭受Na +诱导的离子胁迫。在拟南芥中对 Na+诱导的 Ca2+尖峰缺陷的植物进行的遗传筛选鉴定了编码 MOCA1的基因中的突变。MOCA1是一种葡糖醛酸转移酶,可在质膜上将带负电荷的葡糖醛酸 (GlcA) 基团添加到肌醇磷酸神经酰胺 (IPC) ),产生的糖基肌醇磷酸神经酰胺 (GIPC) 鞘脂可以结合 Na +阳离子,从而诱导细胞膜的去极化。MOCA1 依赖性 GIPC 感知环境 Na +浓度的变化并通过尚未鉴定的 Ca2+转运蛋白产生盐依赖性细胞内 Ca 2+尖峰。两个候选的Ca 2+转运蛋白是AtANN1和AtANN4。除了引发质膜的变化外,高盐度还会通过多种途径破坏细胞壁组织。在拟南芥中,LRX-RALF-FER 模块与感知高盐度诱导的细胞壁破坏有关(见下图 )。在正常条件下,LRX3/4/5蛋白与RALF22/23肽相互作用,防止RALF22/23肽与FER蛋白结合,从而抑制FER蛋白的内化。在盐胁迫下,LRX3/4/5蛋白可以直接感知细胞壁中盐诱导的变化,并且RALF肽与LRX解离以将细胞壁信号转导至FER蛋白,以抑制FER蛋白介导的植物生长途径,启动盐胁迫反应。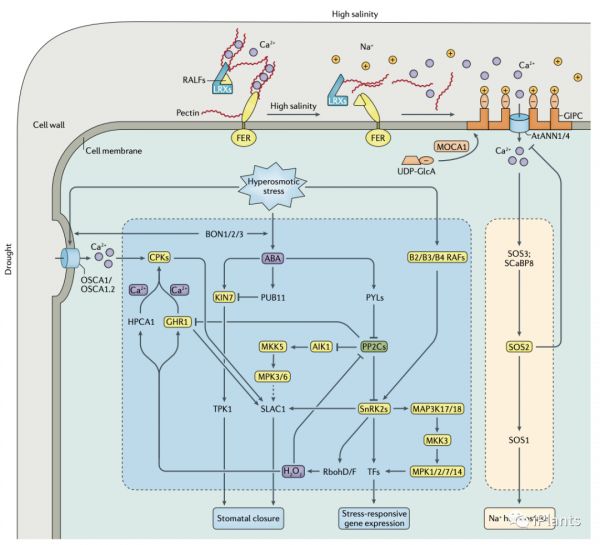
感应温度变化
热胁迫和冷胁迫都可以改变细胞磷脂膜的流动性。冷胁迫的几个假定的传感器已被提出,如膜定位CNGC 离子通道OsCNGC9、OsCNGC14、OsCNGC16和AtANN1通道。鉴于 AtANN1 还介导 NaCl 诱导的 Ca 2+流入,植物对冷胁迫和高盐度的反应是否共享一个调节网络是一个有趣的悬而未决的问题。除了触发细胞质游离钙浓度的瞬时升高外,温度应激还会引起蛋白质稳定性的变化,这意味着可能在所有细胞区室中都感觉到温度应激。例如,在拟南芥中,冷胁迫通过 CBF 转录因子的积累诱导CBF 与 PIF3 相互作用,以减弱 phyB 的降解。冷稳定 phyB 通过控制冷响应基因和生长相关基因的表达来促进冷冻耐受性。此外,拟南芥中的phyB 能感知升高的温度以支持不同的生长模式。因此,phyB 参与植物对温暖和寒冷温度的反应。此外,在拟南芥中,温度升高也可以通过转录阻遏物 ELF3的相变来感知,ELF3 控制着许多参与生长和发育的基因。ELF3 对靶基因的占有率随着温度的升高而降低,导致其表达的激活。虽然细胞膜流动性、蛋白质构象和 RNA 二级结构会受到冷或热应激的影响,但只有热胁迫往往会导致蛋白质变性,并且由此产生的错误折叠蛋白质被某些热休克蛋白 (HSP) 感知。在植物的正常温度下,这些 HSP 结合并阻止热应激转录因子 (HSF) 激活热响应基因的表达并增加各种 HSP 编码基因的转录。然而,在升高的温度下,错误折叠的蛋白质会积累并与 HSP 结合,从而释放 HSF 以激活热应激反应(见下图)。此外,组蛋白变体 H2A.Z 赋予纯化的核小体独特的 DNA 解包特性,也已被证明对拟南芥的温度变化有反应。这些结果导致假设升高的温度导致 H2A.Z 从核小体中解离,这增加了 DNA 对 RNA 聚合酶 Pol II 的可及性,从而导致 HSP 编码基因和其他热响应基因的转录增加。总的来说,这些研究表明,植物不仅可以在细胞表面(包括细胞壁和质膜)感知非生物胁迫,还可以在细胞内区室(如细胞质和细胞核)中感知非生物胁迫。此功能可能允许对细胞内条件进行更直接的响应,并可能赋予定制隔室特定响应的能力。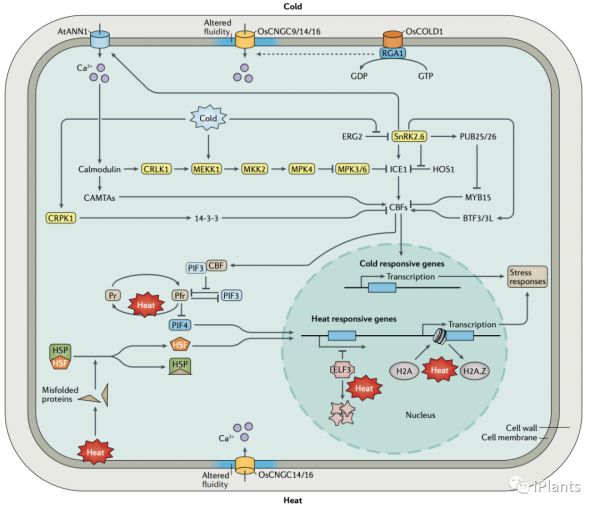
2. 信号转导
对不利环境条件的感知会触发特定于压力的信号转导。该过程涉及第二信使的各种组合,例如 Ca2+、ROS、一氧化氮和磷脂,以及蛋白质的翻译后修饰 (PTM)。该文关注 ROS 信号传导、钙信号传导和蛋白质磷酸化,这是植物响应许多非生物胁迫的信号转导的三个主要特征。Ca2+信号传导
胁迫下Ca2+会通过质膜和细胞器膜流入细胞质,导致细胞质中的 Ca2+峰。在盐胁迫期间,拟南芥中的SOS通路感知到特定的胞质 Ca 2+信号(见上图 2),导致 Na+从根表皮细胞输出到土壤环境,从木质部薄壁细胞到木质部血管,长距离运输到叶子。在 SOS 通路中,SOS3或 SCaBP8 与 SOS2 相互作用并激活 SOS2,SOS2 是 SnRK3 激酶家族(也称为 CIPK)中的一种蛋白质。除了被调节蛋白感知之外,Ca2+信号还可以被转导到钙调蛋白结合转录激活因子 CAMTA1、CAMTA2 和 CAMTA3,它们与CBF基因的启动子结合并激活它们的表达以介导冷应激反应(见下图3 ).蛋白质磷酸化
蛋白质磷酸化是植物对多种非生物胁迫一种常见的和关键的信号转导。如PP2C蛋白磷酸酶家族和 SnRK2 蛋白激酶亚家族的成员是多种应激信号通路的核心参与者。它们调节各种下游蛋白质,包括转录因子,质膜阴离子通道SLAC1控制气孔关闭和质膜NADPH氧化酶RbohF产生细胞外过氧化氢。除了 SnRKs,应激信号转导通常涉及 RLKs 和由 MAPK、MAP2K和 MAP3K 组成的激酶级联反应(如图 3)。ROS信号
植物对各种非生物胁迫作出反应的一个主要特征是产生 ROS,包括超氧阴离子、H2O2、羟基自由基和单线态氧。尽管渗透胁迫下 ROS 的积累可以独立于胁迫诱导的 ABA 积累发生,但 H2O2 的产生显然受 ABA 信号通过 SnRK2 介导的 NADPH 氧化酶 RbohD 和 RbohF 激活的调节。在拟南芥中,细胞外 H2O2可能被富含亮氨酸的重复受体激酶 HPCA1(在整个植物中)和 GHR1(特别是在保卫细胞中)感知以通过激活 Ca 2+通道产生 Ca 2+信号。系统性信号
众所周知,非生物胁迫条件,例如高光和渗透胁迫,会触发植物中的系统性胁迫信号,从而导致未暴露的远端组织中的胁迫反应,从而导致系统性获得性驯化。全株信号同时涉及ROS和Ca2+长距离传输。此外,小肽还可以介导全身应激信号。3. 转录调控
除了通过诱导快速调整来保护植物免受非生物胁迫外,胁迫诱导的信号转导还诱导全基因组转录重编程,从而诱导其他保护机制,如渗透调节、解毒、修复应激引起的损伤和放大或减弱应激信号。应激特异性转录模式通过转录因子连接到上游信号传导。除了胁迫特定的模式,常见的转录反应可以被许多不同的胁迫诱导,如寒冷、高渗和高盐度压力。一般来说,这些常见的应激反应基因编码与 MAPK 级联、钙信号、ROS、细胞凋亡和蛋白质降解相关的蛋白质,并位于不同的细胞区室。ABA 依赖性与 ABA 非依赖性反应
尽管成百上千的基因通过 ABA 信号受到干旱、高盐度和冷胁迫的转录调控,但许多胁迫响应基因是由 ABA 非依赖性机制诱导的(见图 4a)。许多 ABA 调控的基因在其启动子区域包含 ABA 响应顺式元件(ABRE) ACGTGG/TC,而独立于 ABA 的冷响应和干旱响应基因通常包含干旱响应顺式元件(DRE) TACCGACAT。早期与晚期反应
一些基因,特别是那些编码转录因子,显示对胁迫快速早期反应,而有更多的基因是在应激反应后改变,并且由这些早期响应的转录因子驱动的(图 4a中)。在干旱、寒冷或高盐胁迫下的拟南芥中,大多数晚反应基因编码具有保护活性的蛋白质,例如解毒酶和渗透保护剂生物合成。除了加强应激反应外,转录控制还可以导致负反馈调节。例如,在干旱响应过程中,ABA 诱导PP2C的转录基因并抑制它的一些受体基因以衰减ABA信号转导的转录。转录本调控
包括剪接、加帽、聚腺苷酸化和降解在内的 RNA 加工步骤对于植物抗逆性很重要。已经发现 RNA 加工机器的几个组件的功能障碍会严重破坏非生物胁迫抗性,而不会对无胁迫条件下的植物功能产生严重后果。胁迫引起可变剪接
植物在响应于冷和盐胁迫会增加植物的可变剪接,正常剪接的中断会导致功能性蛋白质的产生发生变化,从而改变植物对非生物胁迫或 ABA 的敏感性。如盐胁迫会在拟南芥和木薯 中的几个丝氨酸/精氨酸 (SR) 剪接因子基因中诱导选择性剪接,从而放大剪接调节在应激反应中的作用。胁迫引起的聚腺苷酸化变化
替代性多聚腺苷酸化发生在干旱、热和盐胁迫下。在高粱中,非生物胁迫处理会引起大量基因转录本中 poly(A) 位点选择的变化,从而导致从功能性转录物和翻译产物转变为非功能性转录物。胁迫引起的 microRNA 变化
为应对胁迫条件,植物会调整各种miRNA 的水平。例如,氧化应激转录抑制拟南芥 miR398,由于其目标 mRNA CSD1和CSD2的积累导致氧化应激耐受,这两个基因编码密切相关的 Cu/Zn 超氧化物歧化酶,可解毒超氧化物自由基。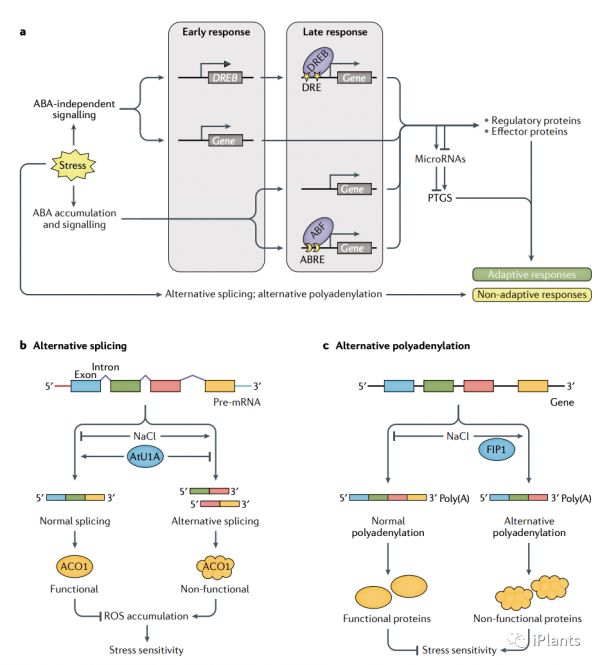
4. 翻译调控
报道的翻译水平的应激反应包括 5' 核糖体暂停、翻译起始的阻断和核糖体的变化。在拟南芥中,热应激引起 5' 核糖体暂停,导致 mRNA 降解,优先影响编码 HSC/HSP70 分子伴侣靶标的转录本(图 5A)。此外,在热应激期间,翻译起始被阻断,编码核糖体蛋白 (RPs) 的 mRNA 优先储存和保护在应激颗粒中;在恢复过程中,这些 mRNA 被释放和翻译,以允许在依赖于伴侣蛋白 HSP101(也称为 CLPB1)的过程中快速恢复翻译。5. 翻译后调控
应激诱导的信号蛋白翻译后修饰
非生物胁迫会显着改变各种类型的 PTM 的水平,这些 PTM 控制蛋白质的定位、积累和/或活性,从而在应激反应的调节中发挥核心作用。PTMs(包括磷酸化)对预先存在的信号蛋白(例如传感器和受体等)的调节以允许其迅速反应,但在应激诱导的转录后从头合成的蛋白质上的 PTMs 也可以重要的。例如,拟南芥磷酸酶 EGR2 通常是 N-肉豆蔻酰化的,这使其能够有效地与 SnRK2.6 相互作用并抑制 SnRK2.6。然而,在冷胁迫下 EGR2 表达增加及其与N-肉豆蔻酰转移酶 NMT1 减弱,导致 N-肉豆蔻酰化 EGR2 被新合成的未肉豆蔻酰化 EGR2 稀释。因此,SnRK2.6 从 EGR2 介导的抑制中释放出来,使其能够调节冷耐受性(图 3)。应激诱导的其他蛋白质的翻译后修饰
PTM 还控制许多对抗压性很重要的非信号蛋白的活性。渗透压力和 ABA 信号导致 SnRK2s 的激活,SnRK2s 磷酸化转录因子、转运蛋白和许多酶,包括参与 ROS 生物发生和清除以及渗透物生物合成的酶。件下正确剪接前 mRNA 所必需的。6. 表观遗传调控
应激反应和应激耐受基因受表观遗传标记调控
已经在许多植物物种中研究表明,表观遗传标记可以响应于各种非生物胁迫。表观遗传变化与参与植物应激反应的基因的转录调控相关联。例如,DNA 甲基化缺陷的拟南芥突变体显示启动子 DNA 低甲基化,因此 Na +转运蛋白基因HKT1 的表达升高因此,增加了耐盐性。胁迫诱导的植物表观遗传模式改变也可能是表观遗传调节因子的变化造成的。例如,番茄果实在冷藏过程中会失去风味,因为冷藏会抑制 DNA 脱甲基酶基因DML2的转录;由此导致负责风味挥发物生物合成的基因启动子处 DNA 甲基化水平的增加导致其表达的沉默(图 5b)。消除胁迫引起的表观遗传变化
认为非生物胁迫诱导的表观遗传修饰可以导致跨代应激记忆是很有吸引力的。然而,在胁迫恢复过程中,存在消除胁迫引起的表观遗传变化的机制。在拟南芥中,染色质重塑蛋白 DDM1 和 DNA 解旋酶 MOM1 通过不同的机制介导表观遗传基因沉默:DDM1 通过 DNA 甲基化起作用,而 MOM1 则没有。热应激诱导的表观遗传沉默的释放不能传递给后代,除非植物同时缺乏 MOM1 和 DDM1 ,这表明这些蛋白质具有冗余功能以防止应激诱导的表观遗传状态的跨代遗传(图 5c)。如果后代没有持续受到胁迫,盐胁迫引起的 DNA 甲基化变化会逐渐消失,这些变化会部分传递给下一代。 因此,植物中胁迫诱导的表观遗传记忆似乎是短暂的,这表明仅通过施加胁迫处理来培育抗胁迫的表观等位基因是不可行的。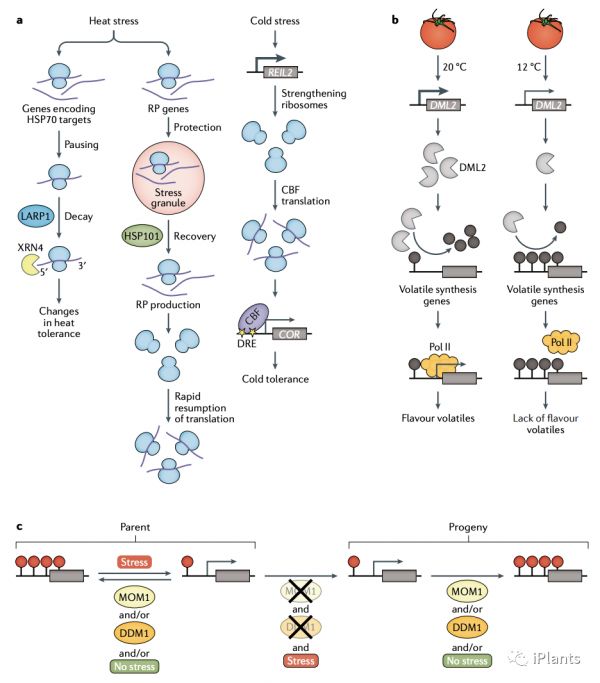
7. 对作物改良的影响
利用作物的自然遗传变异
近年来,通过数量性状基因座方法和全基因组关联研究,越来越多的作物物种中发现了非生物胁迫反应和天然等位基因变异的重要调节因子。通过这些方法鉴定的基因和等位基因可用于培育具有改善的环境适应能力和更高生产力的作物。例如,水稻、小麦 ( Triticum aestivum )和玉米 ( Zea mays ) 中的HKT1等位基因已被确定为控制植物耐盐性的主要数量性状位点,并使得标记辅助育种能够在盐渍土壤中提高小麦产量。. 在非洲水稻亚种Oryza glaberrima 中, TT1 被确定为耐热性的主要数量性状位点,在当地适应中起重要作用。抗逆植物的基因工程
通过上调或下调参与胁迫反应的关键调节因子的表达或活性,可以对抗逆植物进行基因工程改造。尽管可以操纵分子反应的任何级别的调节剂,但大多数成功是通过蛋白激酶和其他信号成分、转录因子、代谢酶和离子转运蛋白实现的。过表达渗透胁迫激活和 ABA 激活的 SnRK2 基因SAPK1和SAPK2导致水稻耐盐性增加,而由 CRISPR-Cas 基因组编辑产生的这些基因的功能丧失突变体显示耐盐性降低,支持重要的SAPK1 和 SAPK2 在这个过程中的作用.基于 CRISPR-Cas 的基因编辑是一种强大的基因工程方法,可用于作物通过碱基编辑、引物编辑和靶向序列插入和替换来产生随机小的 indel 突变或精确的碱基变化。特别是有针对性的序列插入技术使转录或翻译调节序列的高效插入重要的应激反应的基因产生与增加或减少的表达等位基因,使它成为研究和育种的宝贵工具。在不牺牲增长的情况下提高抗压能力
植物应激反应的调节本质上与其他重要的生物过程相协调,尤其是与生长相关的途径。因此,增加作物抗逆性通常伴随着生长减少的不良副作用。然而,在胁迫诱导型启动子控制下表达胁迫耐受基因可以减轻这些生长和产量损失。例如,在水稻中,在干旱诱导型Oshox24启动子的控制下过表达转录因子基因OsNAC6提高了对脱水和高盐胁迫的耐受性,而不会出现组成型过表达观察到的生长迟缓和低谷粒产量。实现胁迫响应和植物生长之间的最佳平衡对于在田间条件下提高产量稳定性的工程作物至关重要。保护植物免受非生物胁迫的化学干预
胁迫抗性也可以通过用影响植物胁迫反应网络分子组分活性的小分子化合物处理来调节。例如,类似 ABA 的活性的化学小分子,包括与 ABA 受体结合、诱导气孔关闭和激活应激反应基因的表达。在植物中使用基因工程胁迫诱导型 ABA 受体可以进一步提高 ABA 模拟物的功效,从而实现更高水平的抗旱性。
结论和未来展望
我们对植物分子对非生物胁迫的反应的理解仍然存在许多差距,特别是在胁迫传感、早期信号、翻译和翻译后调控以及生长调控方面。可能有许多胁迫感受器尚未确定,而大多数已报道的胁迫感受器只能被认为是假设的感受器,因为它们的生理功能和生化传感机制尚不清楚。此外,我们对压力信号的了解是零碎的。大多数已知的信号通路尚未连接到胁迫感受器;对信号通路之间的串扰知之甚少。分子遗传学方法和各种组学方法为我们目前的知识做出了巨大贡献。然而,遗传冗余和致死率,以及分析生物大数据的能力有限,对充分阐明植物应激反应途径和调控网络的尝试构成了重大挑战。基因组编辑工具和人工智能将有助于解决这些问题。此外,需要单细胞组学和其他细胞特异性和组织特异性方法以及原位和实时成像分析方法来了解对非生物胁迫的动态分子反应的时空复杂性。此外,根系微生物群中的许多有益土壤微生物能够增强植物对干旱和其他非生物胁迫的抵抗力,但其潜在的分子机制大多未知。结合遗传、化学和微生物策略的综合方法可以为培育具有高抗逆性和高生产力的植物提供解决方案(图 6)。破译有益微生物保护作用背后的分子机制可能会导致新的策略来提高遗传、化学和微生物方法的功效。相关知识
朱健康
中国健康医疗大数据研究综述——基于期刊论文的分析
Nature综述:饮食调整助力癌症治疗
重磅综述
顶刊综述丨NAT MED(IF:82): 心脏代谢健康、饮食和肠道微生物组: 宏组学视角
Lancet综述:炎症性肠病患者的孕期指南
科学网—抗氧化剂在健康与疾病中的作用机制【自然综述】
综述丨吉林农大: 益生菌对饮食诱导的脂质代谢紊乱的改善作用(国人佳作)
顶刊综述丨NAT REV MICROBIOL (IF:78): 胆汁酸和肠道微生物群: 代谢相互作用和对疾病的影响
健康度假 的翻译是:Health resort 中文翻译英文意思,翻译英语
网址: 【Nature综述】朱健康院士等综述植物非生物胁迫信号转导,为培育抗逆作物提供思路(全文翻译,值得收... https://www.trfsz.com/newsview106563.html
推荐资讯



- 1从出汗看健康 出汗透露你的健 3820
- 2男女激情后不宜做哪些事 3682
- 3早上怎么喝水最健康? 3618
- 4习惯造就健康 影响健康的习惯 3277
- 5五大原因危害女性健康 如何保 3172
- 6连花清瘟、布洛芬等多款感冒药 2948
- 7男子喝水喉咙里像放了刀子一样 2451
- 810人混检核酸几天出结果?1 2216
- 9第二轮新冠疫情要来了?疾控中 2211
- 10转阴多久没有传染性?满足四个 2153
资讯热点排名
资讯热点

