【论肿道麻】Sirtuins: T细胞代谢和功能的重要调节因子
众所周知,代谢是T细胞分化和功能的基础。T细胞代谢活性的变化直接影响其效应功能和命运。因此,了解代谢途径如何影响免疫反应并最终影响疾病进展是至关重要的。表观遗传学和翻译后修饰机制已被发现可以控制免疫反应和代谢重编程。Sirtuins是NAD+依赖的组蛋白去乙酰化酶,在细胞对各种应激的反应中发挥关键作用。最近有报道称其在免疫反应中发挥潜在作用。因此,sirtuins作为治疗免疫相关疾病和增强抗肿瘤免疫的治疗靶点具有重要意义。2022年3月美国 H. Lee Moffitt 肿瘤中心的研究人员于《Experimental & Molecular Medicine》杂志发表了一篇名为 “Sirtuins are crucial regulators of T cell metabolism and functions”的综述,作者综述了Sirtuins在不同亚型T细胞适应性免疫应答过程中的潜在作用。

T细胞介导的适应性免疫反应及其代谢变化 T细胞相关适应性免疫反应在对抗外源病原体的特异性免疫反应中起到至关重要的作用。T细胞在胸腺中发育,成熟后,根据其CD4或CD8受体的表达进行分类。CD4+和CD8+T细胞存在于几个亚群中,这些亚群在免疫应答过程中发挥独特的功能,以特定的表面受体、细胞因子分泌和谱系相关转录因子的表达为特征(图1)。
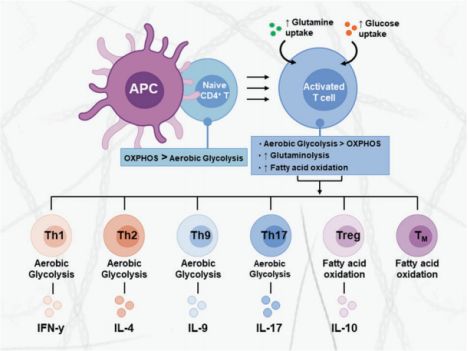
图1. CD4+T细胞亚群的代谢程序
T细胞活化过程启动了多个促进细胞分化和生长的信号通路。细胞从相对静止的氧化代谢转化为强烈的糖酵解代谢,以满足增殖和产生细胞因子的能量需求。每个特异的T细胞亚群的代谢谱被优化,以支持其独特的功能。效应性T细胞(TEFF),主要依赖有氧糖酵解和谷氨酰胺分解来促进其快速生长、增殖和效应功能。Treg细胞主要依赖于脂肪酸氧化 (FA oxidation, FAO)和谷氨酰胺分解来支持其抑制活性。
在初级免疫应答后,一部分CD4+和CD8+T细胞成为记忆T (memory T, TM)细胞,可以对相同抗原作出快速反应。在重新激活前,它们由FAO供能(图1)。肿瘤浸润性T淋巴细胞 (tumor-infiltrating T lymphocytes, TILs)在肿瘤微环境(tumor microenvironment, TME)中代谢受损且功能耗尽,已被证明可以增加FAO代谢作为替代能源。
越来越多的证据表明,免疫细胞的特性、功能和代谢是由错综复杂的信号通路介导的,这些信号通路至少在一定程度上是通过表观遗传学和翻译后修饰(posttranslational modification, PTM) 机制来调控的。Sirtuins是T细胞中关键的表观遗传学和PTM调控因子。以下篇幅将深入阐述Sirtuins如何调控T细胞特异性免疫反应以及Sirtuin调节剂在免疫相关疾病中的治疗潜力。
表观遗传机制 细胞分化主要受表观遗传机制的调控,使DNA序列不变的同时基因表达模式动态变化。基因表达的表观遗传调控发生在DNA、组蛋白和RNA水平上。在这种情况下,DNA甲基化、组蛋白甲基化、乙酰化、泛素化、磷酸化和microRNA依赖的基因沉默已经被很好的阐述。其中重要且常见的机制是组蛋白修饰。组蛋白乙酰转移酶 (histone acetyltransferases, HATs)和组蛋白去乙酰化酶 (histone deacetylases, HDACs)是参与组蛋白乙酰化动态调控的两类酶。乙酰化的组蛋白维持一个开放和流动的染色质结构,称为常染色质,促进基因转录,而去乙酰化的组蛋白通常形成一个紧密排列的染色质结构,阻止基因转录,称为异染色质。
翻译后修饰 PTM是一种生化机制,蛋白质的氨基酸残基在翻译后共价修饰,以调节蛋白质折叠、降解、信号转导、定位、稳定性、酶活性或蛋白质间相互作用。蛋白质乙酰化已成为参与细胞调控的关键PTM,特别是通过修饰组蛋白、核转录调节因子和代谢酶。赖氨酸乙酰化是代谢酶的普遍修饰方式,事实上在人肝组织中几乎所有参与糖酵解、糖异生、三羧酸循环、尿素循环、脂肪酸代谢和糖原代谢的酶都有乙酰化现象。蛋白质乙酰化受到HATs和HDACs之间高度动态平衡状态的调控。 Sirtuins概述 Sirtuin蛋白被归类为III类HDACs,它需要NAD+作为其去乙酰化酶活性的辅助因子。虽然最初被鉴定为HDACs,但后来的研究表明,Sirtuins可以去乙酰化多种非组蛋白。此外,Sirtuins除去乙酰化外还有其他酶功能。在哺乳动物中,有7种Sirtuins (Sirt1-7)广泛表达,具有不同的亚细胞定位和功能。Sirtuins已被阐述可调节多种细胞活动,包括基因组稳定性和细胞代谢,目前因其在介导与炎症、代谢紊乱和癌症相关疾病中的作用而获得广泛关注(图2)。
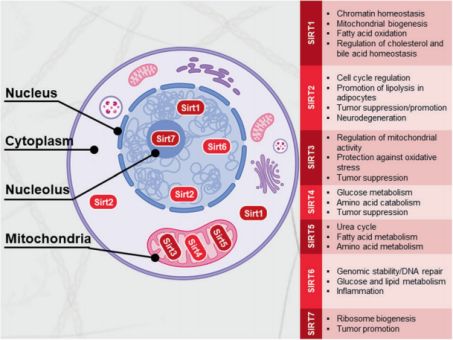
图2. 哺乳动物sirtuins的亚细胞定位和主要功能
SIRT1 Sirt1表达或活性降低有助于T细胞活化,从而导致自身免疫性疾病。此外,Sirt1介导的代谢过程的调节对于优化免疫细胞功能至关重要。
T细胞活化后c-Jun表达上调,诱导IL-2的产生、细胞增殖和分化。事实上,白藜芦醇处理诱导的Sirt1表达和活性的增强阻碍了CD4+T细胞的活化和IFN-γ的产生,进一步证实了Sirt1对Th1分化和IFN-γ分泌的负面影响。动物实验表明,Sirt1−/−T细胞促进增殖和IL-2产生,而Sirt1−/−小鼠更容易发生自身免疫性疾病。进一步的研究表明,IL-2可以通过胞质隔离其上游转录因子FoxO3a来抑制Sirt1的转录,从而逆转T细胞的能量不足。FoxO3a的构成活性形式的表达通过保留Sirt1的表达阻断IL-2介导的T细胞耐受逆转。
B细胞淋巴瘤2相关因子1 (B-cell lymphoma 2-associated factor 1, Bclaf1),被认为是细胞凋亡的启动子。Sirt1可通过去乙酰化Bclaf1启动子区域的组蛋白赖氨酸残基来抑制Bclaf1的活性,导致IL-2基因转录降低。早期小鼠哮喘模型中,抑制Sirt1被证明可减少过敏反应。同时,Sirt1抑制可通过B细胞淋巴瘤/白血病11B(B cell lymphoma/leukemia 11B, Bcl11b)转录激活因子抑制Th2细胞的分化。Bcl11b对于Th2的分化是至关重要的,Sirt1直接与Bcl11b相互作用,并以Bcl11b依赖的方式被招募到启动子模板中,以去乙酰化组蛋白,导致转录抑制。
缺氧诱导因子1-α (hypoxia-inducible factor 1-alpha, HIF-1α)的活性一直与产生促炎细胞因子和限制抗炎细胞因子有关。抑制Sirt1可通过Sirt1-mTOR-HIF-1α轴促进Th9细胞分化和IL-9的产生,Sirt1依赖的糖酵解调控对于指导Th9细胞的分化至关重要。Sirt1对Th17细胞的影响存在争议。Sirt1激活因子,如二甲双胍,已被证明可以阻碍Th17细胞的分化,并通过去乙酰化STAT3转录因子来降低IL-17A和RORγt的表达。另一项研究表明,在体内使用NAD+激活Sirt1延迟了经验性自身免疫性脑脊髓炎(experimental autoimmune encephalomyelitis, EAE)的发生。这种保护作用被认为是因为Sirt1表达增强导致Th17相关炎症反应减少。因此Sirt1抑制剂有预防自身免疫性疾病的潜能。
Foxp3是Treg细胞发育和功能的主要调控因子,Sirt1去乙酰化酶的活性已被发现可以降低Foxp3蛋白水平,而使用Sirt1抑制剂治疗会导致功能性Treg细胞的增加。此外,Sirt1基因缺失或药物抑制通过增加Foxp3mRNA水平,增加Foxp3+Treg细胞的数量和抑制活性。
碱性亮氨酸拉链ATF样转录因子(basic leucine-zipper ATF-like transcription factor, BATF)调节T细胞免疫应答的诸多方面。据报道,Sirt1在BATF的影响下影响CD8+T细胞的分化和效应功能。向FAO的代谢转换是TM细胞分化的特征,由转录共激活因子PGC-1支持,参与线粒体生物合成和线粒体氧化磷酸化(OxPhos)代谢。Sirt1介导的去乙酰化已被证明可以增加PGC-1α和PGC-1β辅助因子的转录活性,这表明Sirt1可能在促进TM细胞形成中发挥作用(图3)。
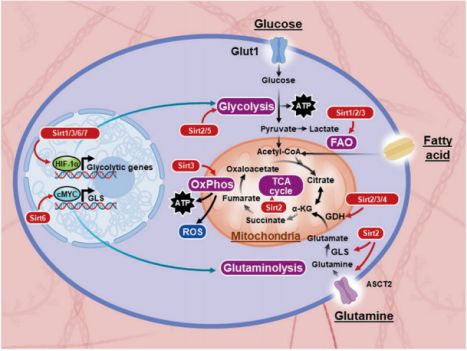
图3. 哺乳动物蛋白和代谢重编程
SIRT2 Sirt2可通过负调控NF-κB p65亚基来发挥抗炎功能。Sirt2缺陷小鼠被发现可通过骨髓来源巨噬细胞的极化发生严重的葡聚糖硫酸钠(dextran sodium sulfate, DSS)诱导的结肠炎,Sirt2−/−小鼠肠系膜淋巴结中活化的CD4+CD69+T细胞比例更大,提示Sirt2在限制CD4+TEFF细胞功能中发挥作用。
最近的一篇文章揭示了Sirt2作为T细胞代谢和功能的主要调节因子的新功能。Hamaidi等人发现,TCR激活后,CD4+和CD8+Sirt2−/−T细胞增殖增加,激活的CD8+T细胞显示IFN-γ产生和颗粒酶B表达增加,导致强大的细胞毒性活性。这种Sirt2−/−T细胞的高反应性表型使小鼠在体内受到肿瘤攻击时具有优越的抗肿瘤免疫力。在T细胞激活过程中,Sirt2缺失可使多种代谢酶高乙酰化后活性增加,导致有氧糖酵解、OxPhos、FAO和谷氨酰胺分解作用增强(图3)。
研究发现,Sirt2在TM细胞阶段表达上调。TM细胞依赖于FAO代谢,Sirt2负向影响FA分解代谢;因此,Sirt2缺失促进体外CD4+和CD8+TM细胞形成,与上游细胞存活和减少凋亡相关。同样,在肿瘤刺激下,Sirt2−/−小鼠的TME和次级淋巴器官中TM细胞增加,而在无瘤WT和Sirt2−/−对照小鼠之间没有观察到定性或定量表型差异。SIRT3 Sirt3主要定位于线粒体基质,在调节线粒体代谢中发挥重要作用,包括TCA循环、尿素循环、FAO和活性氧(reactive oxygen species, ROS)解毒作用(图3)。Sirt3促进能量生成;因此,它在代谢活跃组织中表达更高。
最近的一份报告表明,Sirt3缺陷(Sirt3−/−)小鼠的T细胞导致移植物抗宿主病(graft-versus-host disease, GVHD)的严重程度降低。在该研究中,异体Sirt3−/−T细胞的保护作用与减少T细胞增殖和CXCR3表达有关,但对细胞因子分泌或细胞毒性功能没有显著影响。抑制OxPhos会损害Treg细胞的功能,鉴于Sirt3在OxPhos中的关键作用,预计Sirt3会促进Treg细胞的抑制活性。事实上,Sirt3−/−小鼠的Treg细胞表现出抑制功能受损,这在体外抑制实验和体内同种异体移植模型中得到了证实,HDAC9缺失通过增加Sirt3表达来增加Treg抑制活性。此外,Sirt3也参与了CD8+T细胞的功能。SIRT4 Sirt4是一种具有ADP-核糖基化活性的线粒体sirtuin。Sirt4通过ADP核糖基化谷氨酸脱氢酶(glutamate dehydrogenase, GDH)抑制其酶活性,限制谷氨酰胺转变成谷氨酸并生成ATP(图3)。可以想象,T细胞的代谢活性和效应功能可以通过增加谷氨酰胺分解抑制Sirt4来增强,谷氨酰胺分解是活化T细胞的另一种ATP生成途径。研究发现,Sirt4通过协助重编程人类单核细胞脓毒症模型中的代谢和能量,生理性破坏免疫耐受,并消除急性炎症。
最近的一项动物实验发现,Sirt4在脊髓灰质浸润的Treg细胞中表达上调。Sirt4通过阻断AMPK信号来抑制Treg细胞的抗神经炎症活性,因为AMPK激动剂恢复了Treg细胞中Foxp3和IL-10的表达。然而,这一结果也可能与Sirt4通过谷氨酰胺代谢影响Treg细胞的发育有关。 SIRT5 Sirt5在赖氨酸相关新型酶活性方面的作用是独特的,包括去甲酰化和去戊二酰化。在DSS诱导结肠炎小鼠模型中,已报道Sirt5具有抗炎保护作用。研究发现,Sirt5通过调节丙酮酸激酶M2(pyruvate kinase M2, PKM2)的琥珀酰化来抑制巨噬细胞IL-1β产生和促炎反应。在严重脓毒症中,炎症和免疫抑制状态的诱导和持续通常与继发感染和死亡的风险增加有关。Sirt5被发现可通过促进NF-κB p65乙酰化来缓解内毒素耐受巨噬细胞的炎症反应。最近一项研究表明Sirt5在调节T细胞激活和分化中起着关键作用。
事实上,Sirt5缺失可促进小鼠幼稚T细胞的活化,并增加TCR结合时IFN-γ的产生,并进一步影响T细胞的分化。Sirt5缺失增强了Th1细胞和CTL分化,降低了CD4+ Treg细胞分化,而在稳定状态下,WT和Sirt5−/−小鼠的外周淋巴器官中没有观察到定性或定量的表型差异。更重要的是,尽管Sirt5−/−小鼠对DSS诱导结肠炎高度易感,但Sirt5−/−小鼠对实验诱导结肠炎后结肠肿瘤具有抵抗力,这与结肠组织免疫细胞IFN-γ产生增加有关。值得注意的是,大量脓毒症小鼠模型的研究表明,Sirt5在TEFF细胞功能中作用有限。SIRT6 Sirt6是一种染色质相关的sirtuin,涉及许多生物学功能,包括转录抑制、葡萄糖稳态、DNA修复、端粒功能、细胞分化、有丝分裂和减数分裂。虽然Sirt6具有单ADP糖化酶活性,但其最稳定的活性是组蛋白去乙酰化。
最近的研究表明,Sirt6具有抗炎特性。Sirt6通过影响染色质结构而不是直接去乙酰化NF-κB来抑制促炎基因的转录,而Sirt1和Sirt2直接去乙酰化p65并抑制其转录活性。虽然Sirt6与p65亚基结合,但它通过去乙酰化NF-κB靶基因启动子上的H3K9来调节其转录活性。另外,Sirt6可通过调节不同细胞内信号因子,如Ca2+稳态和mRNA翻译,来诱导促炎症作用。事实上,Sirt6已被证明可以促进细胞因子和趋化因子的表达,并通过增加一些免疫细胞中的mRNA翻译效率来促进TNF分泌。
关于Sirt6在适应性免疫反应中的作用研究较少。它通过抑制基因转录抑制糖酵解。Sirt6已被证明与HIF-1α作用,可通过调节HIF-1α和Myc通路对T细胞代谢和功能具有免疫调节作用(图3)。 SIRT7 Sirt7是sirtuins中最后一个也是研究最少的成员。Sirt7是核糖体生成的重要调控因子,它在核仁中富集,促进RNA聚合酶I依赖的核糖体(ribosomal, r)RNA基因的转录。Sirt7的表达与细胞增殖和致癌活性有关,将Sirt7依赖的核糖体生成调控与细胞周期进程、代谢稳态、应激抵抗、衰老和肿瘤生成联系起来。此外,Sirt7与炎症也有关。 总结 Sirtuins促进抗炎能力已得到充分证明,许多研究小组专注于开发治疗策略,以其在体内的酶活性为靶点。然而,为了达到预期的临床结局,决定是刺激还是阻断sirtuin的活性,以及如何在特定的T细胞亚群中调节特定的sirtuin是至关重要的。由于sirtuins普遍存在,在生物体中功能复杂且部分未知,因此值得进一步研究。
迄今为止,研究人员一直专注于通过促进sirtuins的抗炎活性,治疗多种自身免疫性疾病和其他炎症相关慢性疾病。同时,在抗肿瘤免疫中,通过阻断sirtuins促进T细胞效应活性也被关注。事实上,增强肿瘤反应性T细胞活性在抗癌方面非常有效。阻断T细胞Sirt2和Sirt5具有免疫治疗潜能,可以保护小鼠模型免受结直肠肿瘤发生,说明这种治疗策略预期有效。此外,sirtuins在许多肿瘤模型中有致癌作用,sirtuin抑制剂已被证明可抑制癌细胞,说明靶向sirtuins可能抑制肿瘤增殖,同时提高抗肿瘤免疫。另外,sirtuins可以在TIL扩增或嵌合抗原受体(chimeric antigen receptor, CAR)T细胞体外生成过程中进行基因编辑,从而在免疫细胞治疗时选择性地靶向T细胞。
本文作者还提出,在代谢受限的肿瘤微环境中,肿瘤浸润淋巴细胞耗尽,表现出代谢失调。通过调节sirtuins来增强T细胞的代谢活性,可以为恢复耗尽的肿瘤反应性T细胞提供一种可行策略。
编译:盖晓冬;翁梅琳
审校:张军,缪长虹
参考文献:Hamaidi I, Kim S. Sirtuins are crucial regulators of T cell metabolism and functions. Exp Mol Med. 2022 Mar;54(3):207-215. doi: 10.1038/s12276-022-00739-7. Epub 2022 Mar 16. PMID: 35296782; PMCID: PMC8979958.
声明:古麻今醉公众号为舒医汇旗下,古麻今醉公众号所发表内容之知识产权为舒医汇及主办方、原作者等相关权利人所有。
未经许可,禁止进行转载、摘编、复制、裁切、录制等。经许可授权使用,亦须注明来源。
本文由“健康号”用户上传、授权发布,以上内容(含文字、图片、视频)不代表健康界立场。“健康号”系信息发布平台,仅提供信息存储服务,如有转载、侵权等任何问题,请联系健康界(jkh@hmkx.cn)处理。
相关知识
【论肿道麻】饮食和运动在肿瘤代谢中的作用
疫苗增强CAR-T细胞疗法,或根除某些实体瘤
NK细胞疗法=“肿瘤克星”?NK细胞疗法发展现况大解析
120万元一针癌细胞清零?CAR-T细胞治疗药物是如何发挥作用的
脂肪调节细胞:抑制成脂分化的脂肪干细胞特殊亚群
靶向代谢治疗,推动免疫治疗革命
谷物调节肠道菌群,促进代谢健康
代谢调节
中科西部干细胞研究院:亚健康的干细胞调理机制是什么?
燃脂减肥新方向,首次发现糖原代谢调控脂肪细胞产热和能量消耗
网址: 【论肿道麻】Sirtuins: T细胞代谢和功能的重要调节因子 https://www.trfsz.com/newsview235827.html
推荐资讯




- 1男女激情后不宜做哪些事 4181
- 2从出汗看健康 出汗透露你的健 3839
- 3早上怎么喝水最健康? 3633
- 4习惯造就健康 影响健康的习惯 3283
- 5五大原因危害女性健康 如何保 3187
- 6连花清瘟、布洛芬等多款感冒药 2957
- 7男子喝水喉咙里像放了刀子一样 2455
- 810人混检核酸几天出结果?1 2225
- 9第二轮新冠疫情要来了?疾控中 2219
- 10转阴多久没有传染性?满足四个 2163

