一种生产母乳脂质替代品酿酒酵母菌株及其应用的制作方法
1.本发明涉及一种生产母乳脂质替代品酿酒酵母菌株及其应用,属于生物技术领域。
背景技术:
2.婴幼儿是处于特殊成长阶段的群体,在这一阶段,需要摄入大量且全面的营养,这一时期摄入营养的数量和质量直接影响到婴幼儿未来的生长发育。母乳是婴幼儿获取能量和营养物质的最直接来源,含有大量能够促进婴儿健康生长发育的特殊成分。
3.母乳脂质是母乳的主要营养物质,占母乳的3%-5%,母乳脂质中98%为甘油三酯,是婴幼儿生长发育的主要能量来源。在婴幼儿0-6个月时,母乳脂质提供了婴幼儿所需的40%-50%的能量,同时,母乳脂质还为婴幼儿提供人体必需脂肪酸。研究发现,不仅仅是脂肪酸组成,母乳脂质的脂肪酸分布位置也是关系到婴幼儿是否能吸收母乳中营养成分的关键。母乳脂质中,2号位(sn-2)有将近50%是棕榈酸,这一特定的甘油三酯结构使得婴幼儿能够更好地吸收钙离子,可以有效预防婴幼儿腹泻,避免婴幼儿营养不良,降低婴儿死亡率。然而,母乳喂养虽是婴儿最好的喂养方式,但是在由于某些原因导致无法使用母乳进行喂养的情况下,以母乳为黄金标准的适合婴儿的婴配奶粉就成为替代母乳的理想产品。
4.近年来国内外研究不仅聚焦于母乳脂肪酸组成含量,更加关注于母乳脂肪酸在甘油三酯中的位置分布,在母乳脂肪中,棕榈酸(16:0)酯化到甘油主链的中间(sn-2或β)位置,油酸(18:1)主要酯化到外部(sn-1,3)位置,使三酰甘油(tg)具有独特的立体异构体结构,有助于婴儿肠道的营养吸收。然而,大多数婴儿配方奶粉中使用的脂肪来自动物,与母乳中甘油三酯结构不同的是动物甘油三酯中不饱和脂肪酸如c18:1主要处于甘油主链的中间(sn-2或β)位置,而饱和脂肪酸如c16:0主要处于甘油主链的外部(sn-1,3)位置,这不利于婴儿的吸收与生长。
5.目前,母乳脂质替代品(特定结构的甘油三酯)主要是通过酶法合成,在市场上占据主要地位。如熊志琴(母乳脂替代品的酶法制备及性质研究)以油茶籽油为原料,棕榈酸为酰基供体,在有机溶剂体系中酶法合成sn-2位富含棕榈酸甘油三酯。随着母乳脂质替代品的生产研发,生物从头合成法由于其易于调控,可以合成出人们所需的各种产物愈发受到关注。通过基因工程改造工业生产菌株生产特定脂质组成具有巨大的潜力,同时采用基因工程法构建基因工程菌株从头合成产物其具有低成本、原料不受限制、提取过程简单、无季节性、生产时间短,对环境污染小等诸多优点,从而受到广大学者的青睐。如有研究在拟南芥中使用基因工程合成了一株可以合成母乳替代品的拟南芥植株;在微生物中也有研究者通过改变底物供给合成母乳替代品。但目前尚未有通过从头合成制备母乳脂质替代品的报道。
技术实现要素:
6.为解决上述问题,本发明提供了一种合成母乳脂质替代品的酿酒酵母工程菌,通过将异源的溶血磷脂酸酰基转移酶整合至酿酒酵母以及敲除自身天然溶血磷脂酸酰基转
移酶的方式,增加棕榈酸(c16:0)在酿酒酵母生产的甘油三酯sn-2位置的含量,合成母乳替代脂。并在此基础上敲除代谢途径相关基因,进一步增加产物中母乳脂质替代品的含量。
7.本发明的第一个目的是提供一种生产母乳脂质替代品的酿酒酵母菌株,所述酿酒酵母菌株表达了溶血磷脂酸酰基转移酶crlpaat1,敲除了编码溶血磷脂酸酰基转移酶的基因slc1、ale1和loa1,以及敲除了编码甘油三酯水解酶的基因tgl3、tgl4和tgl5;
8.所述溶血磷脂酸酰基转移酶crlpaat1敲除了自身定位信号肽,并在c端连接了内质网定位信号肽。
9.进一步地,所述溶血磷脂酸酰基转移酶crlpaat1的核苷酸序列如seq id no.4所示。具体地,序列如下:
10.caaaaaagatctcaaaacagaagaaccaacccaagcagccaaaaccaaagcaacaacttgttgagcagcaccagtagcagccaaagctgcagccaaacctctccaagaaccataacaaacaccagcaaaaaccaaaatagaaaccaaagtaatagccaattgttgaacccacatagatgaagaataagaataagcttgtggaccaggcatagaaaatcttggaacatttttcaaatcttccaaaccataagtagcagcccaacaagaatcccaagtagcttgcaaatcagtaaaaaaagcttcaaaatttggataatcagcagatttcaaaaccttagaaaaagtagtaacacaagttctaccaaaatgaacagattgagatttttcagacaaaacttcatctttacctctagtaacaacaatttgaacaggcaattttctagaatgagcataatgcaacatacctcttttcaatggcaaagaagcaggtttagtagatctatgaccttcaggataaaccaacaaaccaggaacatgagaagaacccaaagtttgatccaaccaagcattaaaagcttctttatcagcaatagtacctctcttaaacaaaacaatacctttcaaaatcatgcaagaagtacaaaaaacagggaaaacaaaataaaccaaccatctagacattaaagcagctctaccttcagtcaaataagcatcaataaaaaaatcagcccaagatctatgattacacaaatacaaacatggaccacctttatacaaagtatgttcaccagcttgcaacaaagtaactctaaaataagcaaccaaagctctagcccaatccaacatatcatttcttttacccaaagaagcgaatctgattctatataaaatagcaaagattggtaaagaccaataaaaaacaaaaacagaaaacaagaaagatggtaaacccaaccatttagtcaaaacagacat。
11.进一步地,所述溶血磷脂酸酰基转移酶crlpaat1的自身定位信号肽的核苷酸序列如seq id no.8所示。具体地,序列如下:
12.ttgatccaaccaagcattaaaagcttctttatcagcaatagtacctctcttaaacaaaacaatacctttcaaaatcatgcaagaagtacaaaaaacagggaaaacaaaataaaccaaccatctagacattaaagcagctctaccttcagtcaaataagcatcaataaaaaaatcagcccaagatctatgattacacaaatacaaacatggaccacctttatacaaagtatgttcaccagcttgcaacaaagtaactctaaaataagcaaccaaagctctagcccaatccaacatatcatttcttttacccaaagaagcgaatctgattctatataaaatagcaaagattggtaaagaccaataaaaaacaaaaacagaaaacaagaaagatggtaaacccaaccatttagtcaaaacaga。
13.进一步地,所述内质网定位信号肽为hdel,其核苷酸序列如seq id no.13所示。内质网定位信号肽在异源溶血磷脂酸酰基转移酶中进行表达,对异源酰基转移酶进行修饰,使其可以驻留在内质网中并发挥功能。具体地,序列如下:
14.tcagaacaagcagcacaacaagcagttaataatgcgggctggtcagttatttcagcagcacaactgggctatgcgggcaaaacagatgcaagaggcacatattatggcgaaacagcgggctatacaacagcacaagcagaagttctgggcaaatatgattcagaaggcaatctgacagcaattggcatttcatttagaggcacaagcggcccgagagaatcactgattggcgatacaattggcgatgttattaatgatctgctggcgggcttcggcccgaaaggctatgcagatggctatacactgaaagcatttggccaactgctgggcgatgttgcaaaatttgcacaagcacatggcctgagcggcgaagatgttgtggttagcggccat。
15.进一步地,编码溶血磷脂酸酰基转移酶crlpaat1的基因以质粒pmhylp-leu为表达载体,pmhylp-leu的核苷酸序列如seq id no.5所示。
16.进一步地,所述slc1的核苷酸序列如seq id no.15所示,所述ale1的核苷酸序列如seq id no.16所示,所述loa1的核苷酸序列如seq id no.17所示。
17.进一步地,所述tgl3的核苷酸序列如seq id no.18所示,所述tgl4的核苷酸序列如seq id no.19所示,所述tgl5的核苷酸序列如seq id no.20所示。
18.进一步地,所述重组酿酒酵母菌株以酿酒酵母s.cerevisiae cen pk2-1c、w303、fy1679或by4743为出发菌株。
19.本发明的重组酿酒酵母菌株的构建方法,包括以下步骤,以下步骤可为任意顺序:
20.s1、敲除溶血磷脂酸酰基转移酶基因的自身定位信号肽,并在c端连接内质网定位信号肽,得到去溶血磷脂酸酰基转移酶基因,将所述去溶血磷脂酸酰基转移酶基因导入酿酒酵母中;
21.s2、敲除酿酒酵母自身的溶血磷脂酸酰基转移酶(slc1、ale1和loa1);
22.s3、敲除酿酒酵母自身的甘油三酯水解酶(tgl3、tgl4和tgl5),得到所述重组酿酒酵母菌株。
23.进一步地,在步骤s1中,将溶血磷脂酸酰基转移酶基因采用cre/loxp技术整合至酿酒酵母基因组,包括以下步骤:
24.1)所述溶血磷脂酸酰基转移酶编码基因通过cre/loxp系统如seq id no.5、seq id no.6和seq id no.7所示进行基因整合表达,选用组成型启动子p
tef1
,p
tdh1
,p
pgk1
,p
pyk
,p
ino2
,p
itr1
,p
ald5
,p
ion1
,p
leu2
,p
zwf1
,终止子t
adh1
,t
dnm1
,t
tps1
,t
tdh3
,t
slx5
,t
atp5
,t
cyc1
,设计pcr的引物,使基因表达框相邻片段的重叠区达到40~100bp,构建crlpaat1(ncbi reference sequence:xp_042921325.1)的基因表达整合框。
25.2)通过cre/loxp的方法将溶血磷脂酸酰基转移酶基因的表达框整合至酿酒酵母104c、416d、208c、1622a、308a和911b位点。
26.进一步地,在步骤s2中,使用cre/loxp方法将酿酒酵母自身编码溶血磷脂酸酰基转移酶ale1、loa1和slc1基因敲除。
27.本发明的第二个目的是提供上述酿酒酵母菌株在制备母乳脂质替代品中的应用。
28.进一步地,所述的应用为,以葡萄糖为底物,发酵生产母乳脂质替代品。
29.进一步地,发酵生产过程中,ph为6.0-8.0。
30.进一步地,发酵生产过程中,温度为20-30℃。
31.进一步地,将上述酿酒酵母菌株接种至以葡萄糖为碳源无氨基氮源无菌培养基、限制氮源的ypd无菌培养基、无机盐无菌培养基或限制氮源的大豆蛋白胨无菌培养基中进行发酵,在ph=6.0-8.0,200-300rpm,20-30℃条件下通气发酵。
32.本发明的有益效果:
33.本发明提供的重组酿酒酵母可实现以葡萄糖为碳源的无菌培养基中发酵生产母乳替代脂,为代谢工程改造酿酒酵母合成母乳脂质奠定了基础。本发明提供的重组酿酒酵母的构建方法简单,便于使用,具有很好地应用前景。
附图说明
34.图1为改变异源基因crlpaat1定位;
35.图2为薄层层析分离甘油三酯结果;
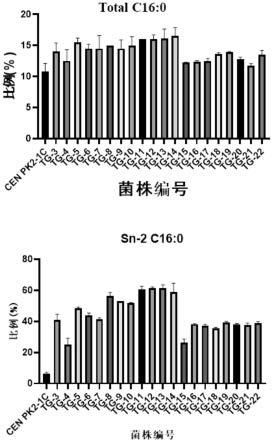
36.图3为不同菌株中总脂肪酸和sn-2位c16:0的相对含量。
具体实施方式
37.下面结合附图和具体实施例对本发明作进一步说明,以使本领域的技术人员可以更好地理解本发明并能予以实施,但所举实施例不作为对本发明的限定。
38.涉及的实验材料:
39.250ml摇瓶,分析天平,灭菌锅,重组菌株,摇床
40.培养基(g/l):
41.ynb培养基:ynb 3.4,硫酸铵10,葡萄糖20,尿嘧啶0.05,组氨酸0.05,亮氨酸0.05,色氨酸0.05;
42.限制氮源的ypd无菌培养基:葡萄糖20,酵母提取物10,蛋白胨10;
43.无机盐无菌培养基:葡萄糖10,(nh4)2so
4 1,k2hpo
4 0.125,khpo
4 0.875,ki 0.0001,mgso4·
7h2o 0.5,cacl2·
2h2o 0.l,nacl 0.1,维量元素母液lml,维生素母液lml;
44.以及限制氮源基础发酵培养基:葡萄糖20,硫酸铵4,kh2po
4 2.5,mgso4·
7h2o 0.5;
45.种子培养基:ynb培养基,无机盐无菌培养基,限制氮源的ypd无菌培养基,限制氮源基础发酵培养基;
46.发酵培养基:ynb培养基,无机盐无菌培养基,限制氮源的ypd无菌培养基,限制氮源基础发酵培养基。
47.涉及的检测方法:
48.气相色谱-质谱联用仪(gc-ms)检测法:岛津(gcms-qp2010 se),sh-rtx-wax色谱柱,流动相为he,柱温100℃,进样量1μl,分流比20:1,流量1.0ml/min。
49.本发明中,以重组酿酒酵母s.cerevisiae cen pk2-1c mata;ura3-52;trp1-289;leu2-3,112;his3-δ1;mal2-8c;suc2;s.cerevisiae w303 mata;ura3-1;trp1-δ1;leu2-3,112;his3-11;ade2-1;can1-100;s.cerevisiae fy1679 mata;ura3-52;trp1-δ63;leu2-δ1;his3-δ200;gal2和s.cerevisiae by4743 mata;ura3-δ0;met15-δ0;leu2-δ0;his3-δ1;lys-δ0为出发菌株,采用cre/loxp方法将衣藻属来源的溶血磷脂酸酰基转移酶整合至酿酒酿酒酵母基因组,获得具有合成母乳脂质替代品(甘油三酯)能力的酿酒酵母菌株。
50.酿酒酵母是又称面包酵母或者出芽酵母。酿酒酵母是与人类关系最广泛的一种酵母,作为食品安全菌株已用于制作面包和馒头等食品、酿酒工业。近年来,学者们开始研究利用酿酒酵母生产天然产物,如青蒿酸、三七皂苷等。酿酒酵母具有高安全性,低致病性,高抗逆性,而且受噬菌体污染的概率较低等优点,因此它也在基因工程领域发挥着重要作用。然而,酿酒酵母自身甘油三酯结构与母乳相差较大。因此,为了获得母乳脂质替代品生产工程菌株,我们采用cre/loxp技术将酰基转移酶代谢途径整合至酿酒酵母,以达到生产母乳脂质替代品的效果。
51.以下实施例所用引物见下表:
[0052][0053]
[0054][0055]
实施例1异源酰基转移酶表达酿酒酵母菌株(tg-1~tg-4)的构建
[0056]
根据ncbi上公布的衣藻属来源的溶血磷脂酸酰基转移酶crlpaat1(ncbi reference sequence:xp_042921325.1),按照酿酒酵母密码子偏好性进行密码子优化并进行全基因合成。根据重叠衍生pcr引物设计方法,设计引物使基因表达框相邻片段的重叠区达到40~100bp。设计引物扩增酿酒酵母cen pk2-1c染色体911b位点的上下游同源臂,启动子p
tef1
,p
tdh1
,p
pgk1
,p
pyk
,p
ino2
,p
itr1
,p
ald5
,p
ion1
,p
leu2
和p
zwf1
,终止子t
adh1
,t
dnm1
,t
tps1
,t
tdh3
,t
slx5
,t
atp5
和t
cyc1
和信号肽(hdel)片段。
[0057]
以全基因合成的crlpaat1质粒为模板,设计引物异源溶血磷脂酸酰基转移酶。以质粒pmhylp-leu为模板(如seq id no.5所示)进行pcr,扩增获得缺陷型标签片段,通过重叠延伸pcr获得基因整合框。
[0058]
(1)通过引物f1和r1;f2和r2扩增得到911b位点的上下游同源臂片段,通过引物f3和r3扩增得到启动子p
tef1
片段,通过引物f4和r4扩增得到终止子t
adh1
片段,通过引物f5和r5扩增得到seq id no.4所示的crlpaat1基因片段,通过引物f6和r6扩增获得pmhylp-leu片段,通过重叠延伸pcr构建基因表达框并将其转入酿酒酵母cen pk2-1c,获得菌株tg-1。
[0059]
(2)使用f5和r7将crlpaat1基因(基因序列如seq id no.4所示)自身的定位信号肽(如seq id no.8所示)进行敲除,获得mcrlpaat1基因,将(1)中的同源臂、(f7和r3)启动子、终止子、缺陷型标签片段按照(1)的方法构成的基因表达框转入酿酒酵母cen pk2-1c中,获得菌株tg-2。
[0060]
(3)通过引物f8和r8,f9和r9扩增得到与mcrlpaat1基因的c端和n端相连的信号肽hdel(如seq id no.13所示)片段,将与mcrlpaat1基因c端连接的信号肽hdel的片段与(2)中的片段构成基因表达框转入酿酒酵母cen pk2-1c中获得菌株tg-3,将与mcrlpaat1基因n
端连接的信号肽hdel的片段与(2)中的片段构成基因表达框转入酿酒酵母cen pk2-1c中获得菌株tg-4。
[0061]
对tg-3和tg-4菌株进行检测,发现tg-3菌株tag的sn-2位置的c16:0的相对含量增加至40%,因此选择tg-3菌株进行后续实验。以上过程均通过醋酸锂转化方法将构建好的基因整合框转化至酿酒酵母感受态细胞,挑取菌落进行pcr验证并选取部分pcr正确的转化子进行测序验证。
[0062]
实施例2高产母乳脂质替代品重组酿酒酵母菌株(tg-5~tg-14)的构建
[0063]
根据重叠衍生pcr引物设计方法,以酿酒酵母cen pk2-1c基因组为模板,扩增获得(f10,r10和f11,r11)slc1、(f12,r12和f13,r13)ale1和(f14,r14和f15,r15)loa1基因上下游同源臂,以质粒(f16,r16)pmhylp-leu、(f17,r17)pmhylp-his和(f18,r18)pmhylp-trp为模板(如seq id no.5~seq id no.7所示)进行pcr,扩增获得缺陷型表达框片段。通过重叠延伸pcr获得基因整合框并在tg-3菌株的基础上,分别对上述三个基因进行单敲除、双敲除和三敲除,获得tg-5(敲除slc1)、tg-6(敲除ale1)、tg-7(敲除loa1)、tg-8(敲除slc1、ale1)、tg-9(敲除slc1、loa1)、tg-10(敲除ale1、loa1)和tg-11(敲除slc1、ale1、loa1)菌株。
[0064]
对tg-5~tg-11菌株的总脂肪酸和sn-2位c16:0的相对含量进行检测,结果见图3,其中,菌株tg-11总脂肪酸中c16:0的含量最高,且sn-2位c16:0的相对含量达到了60%以上。
[0065]
在tg-11菌株的基础上对酿酒酵母甘油三酯水解途径的相关基因进行敲除,使用之前的方法进行引物设计,扩增得到tgl3(f19,r19和f20,r20)、tgl4(f21,r21和f22,r22)和tgl5(f23,r23和f24,r24)基因上下游同源臂和缺陷型表达框(f25~f27和r25~r27)片段,依次将这三个基因敲除表达框转入tg-11菌株中获得tg-12(敲除tgl3)、tg-13(敲除tgl3、tgl4)和tg-14(敲除tgl3、tgl4、tgl5)菌株。
[0066]
对tg-12~tg-14菌株的总脂肪酸和sn-2位c16:0的相对含量进行检测,结果见图3,最终检测得到tg-14菌株的总脂肪酸含量相对于tg-11有所提升,且tag的sn-2位置的c16:0的相对含量增加至60%。以上过程均通过醋酸锂转化方法将构建好的基因整合框转化至酿酒酵母感受态细胞,挑取菌落进行pcr验证并选取部分pcr正确的转化子进行测序验证。
[0067]
实施例3重组酿酒酵母发酵生产母乳脂质替代品
[0068]
本发明中所有的重组酿酒酵母菌株均通过以下方式进行发酵。将重组的酿酒酵母重组菌株划线与无氨基氮源平板(缺乏缺陷型相对应的氨基酸),30℃下培养直至长出大量菌落。
[0069]
挑取单菌落至种子培养基(选用以葡萄糖为碳源的无氨基氮源无菌培养基、限制氮源的ypd无菌培养基、无机盐无菌培养基以及限制氮源的大豆蛋白胨无菌培养基的任一培养基作为发酵培养基均可),在30℃,220rpm培养18~20h至细胞生长对数期。
[0070]
按起始2~5%的接种量将种子培养液接种至发酵培养基中(选用以葡萄糖为碳源的无氨基氮源无菌培养基、限制氮源的ypd无菌培养基、无机盐无菌培养基以及限制氮源的大豆蛋白胨无菌培养基的任一培养基作为发酵培养基均可),30℃220rpm培养72h。72h后停止培养,将发酵结束的酵母细胞离心之后冻干,用于后续的检测分析。
[0071]
实施例4重组酿酒酵母母乳脂质替代品提取与检测
[0072]
本发明中所有的重组酿酒酵母菌株中的脂质均通过以下方式进行提取。在冻干的
菌体中加入十五烷酸甘油三酯作为内标,加入甲醇和玻璃珠进行振荡破碎,然后将破碎完的溶液转移到新的容量瓶中,加入氯仿,将溶液置于超声水浴10分钟,将超声完成的上清转移至新的容量瓶中,再次加入1.5ml甲醇/氯仿溶剂进行提取,重复该步骤两次,并将所得的脂质提取物进行合并,在合并的溶液中加入2.5ml氯仿和3ml nacl水溶液。剧烈振荡样品,10000rpm离心5min,弃上层液体,将下层有机相转移至新的玻璃管中,使用氮吹仪吹干有机相,使用正己烷进行复溶。使用薄层层析方法对提取的脂质进行分离,展层剂为正己烷:乙醚:乙酸(70:30:1,v:v:v),使用溴百里酚蓝对层析版上的脂质进行显色,根据显色结果使用取样器将甘油三酯条带取下,然后将分离得到的甘油三酯样品置于10ml离心管中,再向试管中加入0.2ml正己烷。之后加入50mg胰甘油三酯水解酶和2ml tris-hcl(ph=8)缓冲液,小心摇动,之后再加入0.5ml胆酸钠溶液,0.2ml氯化钙溶液,盖上离心管盖,小心摇动,随后将离心管置于40℃的水浴锅中5min,期间保持手摇,水浴后去除振荡器上振荡2min,之后加入1ml盐酸溶液(6mol/l)和1ml乙醚,盖上盖子,剧烈震荡10s,在4000rpm离心4min,吸取有机相,氮吹至200ul后使用gc-ms进行测定。基因定位结果如图1,薄层结果如图2,c16:0含量占比如图3。
[0073]
从图1中可看出,将异源基因crlpaat1直接整合在酿酒酵母基因组时,基因的表达量较低;将异源基因自身信号肽进行敲除之后得到的mcrlpaat1基因,在酿酒酵母中表达量明显提高,同时可以发现基因主要在胞质中游离表达;在mcrlpaat1基因c端添加信号肽之后得到rmcrlpaat1基因,该基因在酿酒酵母中表达较强,且在内质网中定位。
[0074]
对比例1
[0075]
将溶血磷脂酸酰基转移酶替换为以下序列(1),敲除自身的定位信号肽(序列如seq id no.22所示),并在敲除后基因的c端添加信号肽hdel,具体步骤同实施例1,得到菌株tg-15;
[0076]
将溶血磷脂酸酰基转移酶替换为以下序列(2),在基因的c端添加信号肽hdel,具体步骤同实施例1,分别得到菌株tg-16、tg-17。
[0077]
(1)芸苔属来源的溶血磷脂酸酰基转移酶lpat1(ncbi reference sequence:af111161),基因序列如seq id no.1所示;
[0078]
(2)智人来源的溶血磷脂酸酰基转移酶agpat1(ncbi reference sequence:nc_000006.12)和agpat2(ncbi reference sequence:cah71722.1),基因序列分别如seq id no.2和3所示;
[0079]
对比例2
[0080]
将信号肽hdel替换为以下信号肽:(1)srp14(基因序列如seq id no.9所示)、(2)srp54(基因序列如seq id no.10所示)、(3)cyb5(基因序列如seq id no.11所示)、(4)sec12(基因序列如seq id no.12所示)、(5)fehdel(基因序列如seq id no.14所示),具体步骤同实施例1的菌株tg-3,分别得到菌株tg-18~tg-22。
[0081]
显然,上述实施例仅仅是为清楚地说明所作的举例,并非对实施方式的限定。对于所属领域的普通技术人员来说,在上述说明的基础上还可以做出其它不同形式变化或变动。这里无需也无法对所有的实施方式予以穷举。而由此所引申出的显而易见的变化或变动仍处于本发明创造的保护范围之中。
相关知识
世界母乳喂养周:母乳代用品可以替代母乳吗?
盘点14株国内婴幼儿食品可用菌株—附法规依据
酵母与肠胃健康:一种微妙的共生关系
微观盟友:揭秘益生菌的健康密码与应用前景
9种最好的益生菌,根据你的需要
益生菌的作用这么多,还有哪些临床应用是你不知道的?
肠道菌群与健康:探究发酵食品、饮食方式、益生菌和后生元的影响
发酵乳调控人体肠道营养健康的研究进展
【安琪酵母】福邦有机肥,营养,使用范围广,提高作物品质!
四大发酵食品的瘦身秘诀
网址: 一种生产母乳脂质替代品酿酒酵母菌株及其应用的制作方法 https://www.trfsz.com/newsview339616.html
推荐资讯




- 1从出汗看健康 出汗透露你的健 3820
- 2男女激情后不宜做哪些事 3682
- 3早上怎么喝水最健康? 3618
- 4习惯造就健康 影响健康的习惯 3277
- 5五大原因危害女性健康 如何保 3172
- 6连花清瘟、布洛芬等多款感冒药 2948
- 7男子喝水喉咙里像放了刀子一样 2451
- 810人混检核酸几天出结果?1 2216
- 9第二轮新冠疫情要来了?疾控中 2211
- 10转阴多久没有传染性?满足四个 2153

