医学1区if18+
肝脂肪变性是最常见的慢性肝病之一,在过去几十年里,全球肝脂肪变性的患病率和发病率一直在上升。肝脂肪变性的发病机制复杂且尚不完全清楚,但胰岛素抵抗(IR)被认为是其发展的主要致病因素。在生理状态下,胰岛素抑制肝脏葡萄糖异生,促进脂肪从头合成(DNL)以储存热量。在IR期间,胰岛素不能抑制肝脏葡萄糖的产生,肝脏DNL仍无限制地增加,胰岛素水平与肝脏脂质沉积之间也存在正相关关系,进而造成肝脂肪变性增加。但肝脂肪变性通过何种机制增加仍未完全明确。
2023年9月21日,山东第一医科大学附属省立医院赵家军、薄涛、高聆共同通讯在Nature Metabolism发表了题为“Upregulation of WDR6 Drives Hepatic de novo Lipogenesis in Insulin Resistance in Mice”的研究论文。发现WD重复域6(WD repeats domain 6,WDR6)促进胰岛素抵抗期间肝脏DNL。并从中药绞股蓝中筛选和转化了一个天然小分子化合物—XLIX。该化合物可靶向WDR6,抑制其介导的肝脏脂质合成,为预防和治疗NAFLD提供了新的潜在治疗靶点和策略。

摘要
正常情况下,胰岛素促进肝脏新生脂肪生成(DNL)。然而,在胰岛素抵抗(IR)期间,当胰岛素信号减弱并伴有高胰岛素血症时,肝脏DNL的促进作用持续不减,肝脏脂肪变性增加。在本研究中,我们发现WD40重复蛋白6 (WDR6)在IR过程中促进肝脏DNL。机制上,WDR6与丝氨酸/苏氨酸蛋白磷酸酶1 (PPP1CB)的β型催化亚基相互作用,促进PPP1CB在Thr316位点的去磷酸化,进而通过dna依赖性蛋白激酶和上游刺激因子1增强脂肪酸合成酶的转录。通过分子动力学模拟分析,我们发现了一个小的天然化合物XLIX,它可以抑制WDR6与PPP1CB的相互作用,从而降低IR态的DNL。总之,这些结果揭示了WDR6是治疗肝脂肪变性的一个有前景的靶点。
1.在胰岛素抵抗期间,WDR6在胰岛素的作用下上调
通过给动物喂食HFD 6周建立了IR小鼠模型。随后进行胰岛素注射(图1a)。AKT(胰岛素信号通路的关键读数)的磷酸化显示对胰岛素的应答降低,表明对胰岛素有抵抗(图1b)。然而,在HFD小鼠中,FASN继续表现出对胰岛素的相同反应(即升高)。结果表明,胰岛素在IR期间仍然可以驱动该脂肪生成基因的表达。为了确定在IR状态下仍对胰岛素有应答的基因,在胰岛素注射和溶剂注射的IR小鼠中进行了肝脏转录组分析。总共确定了263个差异表达基因(DEGs)(图1c)。为了缩小感兴趣的候选基因数量,利用了公共基因表达谱数据集GSE66676和GSE48452,这两个数据集分别是成人和青少年非酒精性脂肪性肝病(NAFLD)/非酒精性脂肪性肝炎(NASH)的微阵列数据库。在三个数据库中,共发现310个基因相交,并与FASN水平呈正相关(图1c)。在这些基因中,根据转录组分析的DEGs,重点关注那些在IR期间对胰岛素产生反应的基因。最终筛选出8个候选基因(图1c)。在注射胰岛素的IR小鼠中进行逆转录定量PCR (RT-qPCR)(图1d),并结合文献调查以确定与胰岛素信号传导和/或代谢相关的基因。据报道,WDR6, 8个候选基因之一,在胰岛素样生长因子-1 (IGF-1)或胰岛素处理的小鼠下丘脑来源的GT1-7细胞中增加。此外,在HeLa细胞中,WDR6可以与LKB1相互作用,LKB1是一个关键的丝氨酸/苏氨酸激酶,通过激活ampk调节能量代谢。提示WDR6可能对胰岛素有应答并参与代谢。最后,我们选择了Wdr6进行进一步研究。
直接检测胰岛素波动的IR小鼠肝脏中WDR6蛋白水平进一步验证了这一发现。胰岛素注射或6小时再喂养后,WDR6水平升高,这分别对应于外源性或内源性胰岛素水平升高的情况(图1e)。此外,发现NAFLD小鼠的肝脏WDR6表达增加,这支持WDR6参与肝脏脂质代谢的假设(图1f)。这些体内外实验结果表明,在IR中,肝脏Wdr6通过INSR信号通路对胰岛素产生特异性反应。
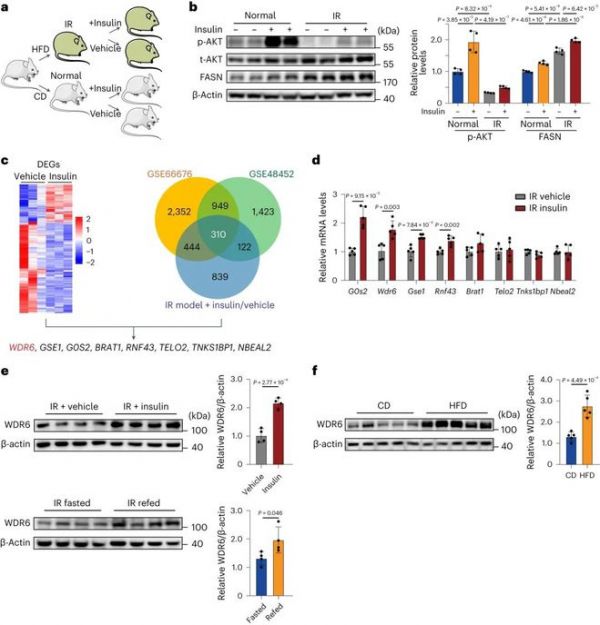
图1:在胰岛素抵抗状态下,WDR6对胰岛素应答而上调
2.在胰岛素抵抗中,Wdr6缺失可防止肝脂肪变性
为了验证WDR6是否参与ir诱导的肝脏代谢紊乱,通过腺相关病毒(AAV)介导的WDR6 shRNA (HBAAV2/8-TBG-micro-mWdr6;shWDR6老鼠)。对照组小鼠注射对照shRNA (shNC小鼠)。在肝脏中,超声分析、油红O (ORO)染色、透射电子显微镜(TEM)和TAG含量显示,与shNC组相比,雄性shWDR6小鼠肝脏脂质沉积减轻(图2a-d)。提示WDR6参与肝脏脂质代谢。为了明确靶向肝脏WDR6是否可以改善HFD的不良反应,将AAV-shWDR6注射到已经喂食了2周HFD的10周龄小鼠中(因为aav介导的表达需要约3-5周才能起作用,注射开始于HFD开始后2周),以模拟靶向WDR6的治疗效果(TM- shwdr6组)。对照组小鼠(注射AAV-shNC 2周的高脂饮食)分为两个亚组,一个亚组在注射AAV-shNC后继续高脂饮食(TM-shNC组),另一个亚组转换为普通饮食。与TM-shNC组相比,TM-shWDR6组hfd诱导的IR和肝脏脂质沉积减轻(图2e,f)。
还通过CRISPR-Cas9技术创建了肝脏特异性敲除Wdr6 (Wdr6 - lko)小鼠。肝脏中TAG沉积减少(图2g,h),磁共振成像也观察到较低程度的脂肪肝(MRI;图2 i, j)。这些结果表明,靶向肝脏中的Wdr6可减少hfd相关的肝脏脂质沉积。综上所述,这些数据表明,肝脏特异性抑制Wdr6可改善或逆转IR期间的代谢紊乱,尤其是肝脏脂质沉积,而Wdr6过表达则有相反的作用。
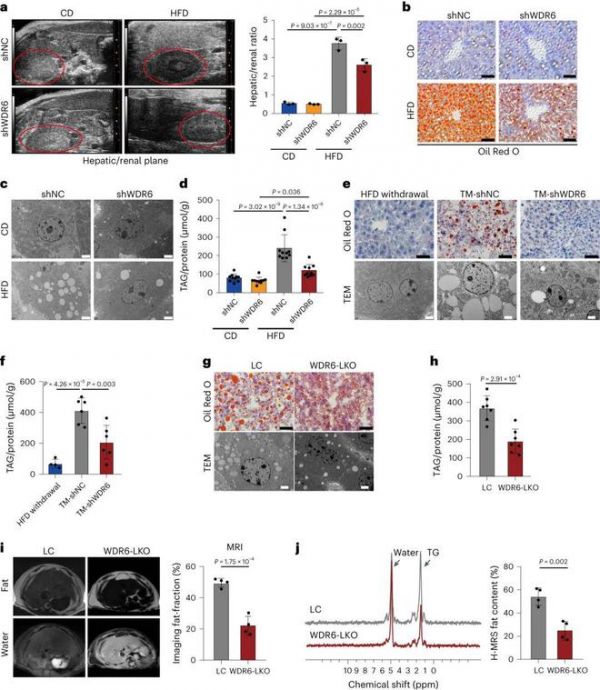
图2:肝脏特异性干预Wdr6对高脂饮食诱导的胰岛素抵抗期间肝脏过度三酰甘油沉积有保护作用
3.肝脏WDR6通过调节新生脂肪生成影响肝脏三酰甘油沉积
然后,我们分别对cd喂养组和hfd喂养组的shWDR6和shNC小鼠的肝脏进行了脂质组学分析,以了解可能受WDR6影响的脂质代谢途径。如图3a和Extended Data Fig. 6a所示,hfd喂养组之间差异最大的顶级代谢物是TAG。在shWDR6和shNC小鼠中,我们评估了参与肝脏TAG代谢的主要通路,包括DNL、脂肪分解、游离脂肪酸氧化和摄取的各种代表性基因,以及一些葡萄糖代谢的代表性基因的表达模式。其中,与shNC小鼠相比,shWDR6小鼠dnl相关基因Fasn在mRNA和蛋白水平均降低。葡萄糖代谢基因的转录在组间没有明显的差异(图3b和Extended Data Fig. 6b,c)。此外,在HFD组中,与对照小鼠相比,Wdr6敲低后FASN的酶活性也较低(图3c)。
为了进一步证实我们的发现,进行了碳同位素标记分析,以估计在Wdr6全身敲除小鼠(Wdr6 - wko)和野生型对照的原代肝细胞中新合成的棕榈酸的含量。来自示踪剂D-[统一标记的13C (U-13C)]-葡萄糖的13C标记的棕榈酸盐在WDR6-WKO肝细胞中显著低于WT(图3d)。此外,肝脏DNL发生率通过使用[11C]乙酸盐正电子发射断层扫描(PET)进行估计,肝脏Wdr6敲低降低了DNL发生率(图3e)。另外,由于TAG的侧链由各种各样的FAs组成,不同的生物合成酶参与,我们发现差异最明显的FAs是饱和的(FA16:0, FA18:0)和单不饱和的(FA16:1, FA18:1),分别是FASN或SCD1的直接或近端产物(Fig. 3f)。综上所述,这些结果表明WDR6在IR过程中促进肝脏DNL,其中Fasn可能是一个主要的作用靶点。接下来,我们测试了WDR6是否可以介导胰岛素在IR期间调节FASN的过程。HFD-shWDR6小鼠注射胰岛素,6小时后安乐死。与HFD-shNC小鼠不同,敲低Wdr6显著减弱了胰岛素增加FASN水平的能力,但对ACC1或SCD1的影响很小(图3g),这表明在IR期间,Wdr6上调可作为胰岛素增加FASN水平和DNL的另一种手段。
接下来,我们想知道wdr6介导的Fasn表达调控是否依赖于DNL7的关键转录因子SREBP1c。在srebp1c基因敲除的C57BL6小鼠中过表达WDR6。虽然SREBP1c缺失可减少肝脏TAG沉积(图3h,i),但与之前报道的结果相似,WDR6过表达仍可增强SREBP1c- ko小鼠肝脏TAG和FASN蛋白水平(图3h - j)。这些结果表明,SREBP1c对于wdr6介导的Fasn表达调控和肝脏TAG沉积的作用是重要的,但不是必需的,因为删除SREBP1c只能部分阻断这种作用。
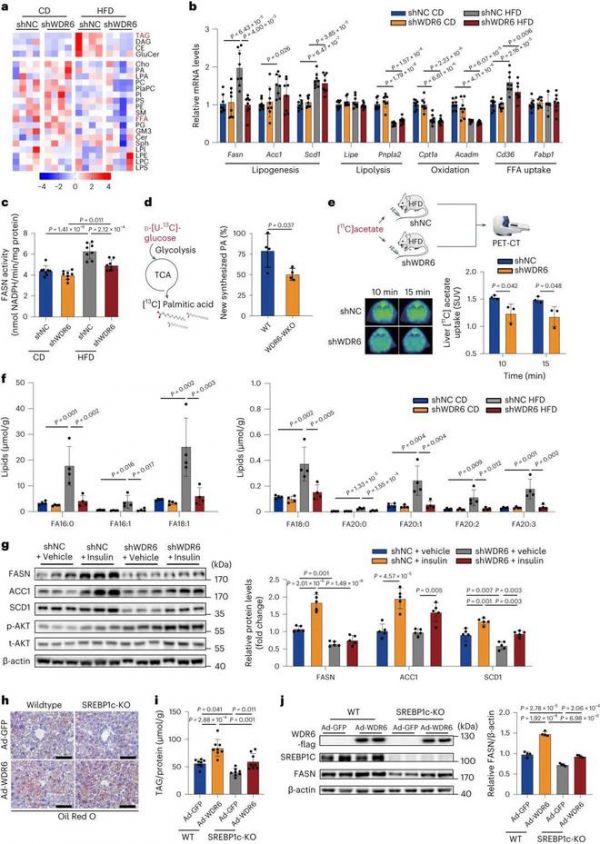
图3:肝脏WDR6通过调节新生脂肪生成影响三酰甘油沉积
4.WDR6与PPP1CB在Thr316相互作用并使其去磷酸化
继续探索WDR6调控FASN表达的机制。选择对WDR6 - FLAG细胞进行FLAG亲和力免疫沉淀(IP)(扩展数据图1),以确定WDR6的结合伴侣(图4a)。选择SDS-PAGE观察到的优势条带进行质谱鉴定(MS;图4 b)。在IP-MS分析的同时,我们还对来自WDR6-WKO和WT小鼠的原代肝细胞进行了磷酸化蛋白质组学分析(图4a)。蛋白质组学分析是作为内部对照进行的。与对照组相比,在WDR6-WKO原代肝细胞中,磷酸化thr316的PPP1CB)表达增加(图4c)。此外,在WDR6 - flag IP洗脱样品中也发现了PPP1CB,确定其是WDR6的结合伴侣(图4b)。因此,PPP1CB出现在两个不同的数据集中(图4a)。这些结果促使我们假设Wdr6缺乏与磷酸化PPP1CB (Thr316)增加相关,这可能是因为Wdr6是PPP1CB的直接或间接伙伴。为了进一步验证WDR6和PPP1CB的相互作用,我们对WDR6 - flag细胞进行了IP分析。我们发现内源性WDR6与PPP1CB相互作用,但与PPP1CA或PPP1CC没有明显的相互作用(图4 d)。此外,在HEK293细胞中,重组WDR6-FLAG和PPP1CB-HA,或重组HA-WDR6和PPP1CB-FLAG共过表达,以进行相互的FLAG亲和力IP分析。我们发现,在这两种情况下,WDR6和PPP1CB之间都存在(直接或间接)物理相互作用(图4e、f)。
接下来,研究了磷酸化ppp1cb (Thr316)是否受到WDR6的影响。检测磷酸化ppp1cb (Thr316)在Wdr6肝特异性敲入和WDR6-LKO小鼠,使用特异性识别Thr316磷酸化ppp1cb的定制一抗。与相应的WT同窝小鼠相比,WDR6-LKI小鼠的肝脏磷酸化ppp1cb (Thr316)水平较低(图4g),相反,WDR6-LKO小鼠的肝脏磷酸化ppp1cb (Thr316)水平较高(图4h)。同时检测磷酸化ppp1cb (Thr316)对外源性/内源性胰岛素的反应。我们发现,与正常对照相比,在hfd诱导的IR小鼠中,磷酸化ppp1cb (Thr316)水平较低(图4i),在胰岛素治疗6小时的IR小鼠中(0.75 U / kg体重),与IR对照组小鼠相比(图4j)较低。此外,与正常喂养相比,空腹16小时(胰岛素水平较低)后磷酸化ppp1cb (Thr316)水平升高,在6小时再喂养(胰岛素水平升高;图4 k)。上述处理的FASN水平均出现了预期的相应变化。这些结果表明,磷酸化的ppp1cb (Thr316)受到WDR6的影响,以及胰岛素和外部营养环境的影响。
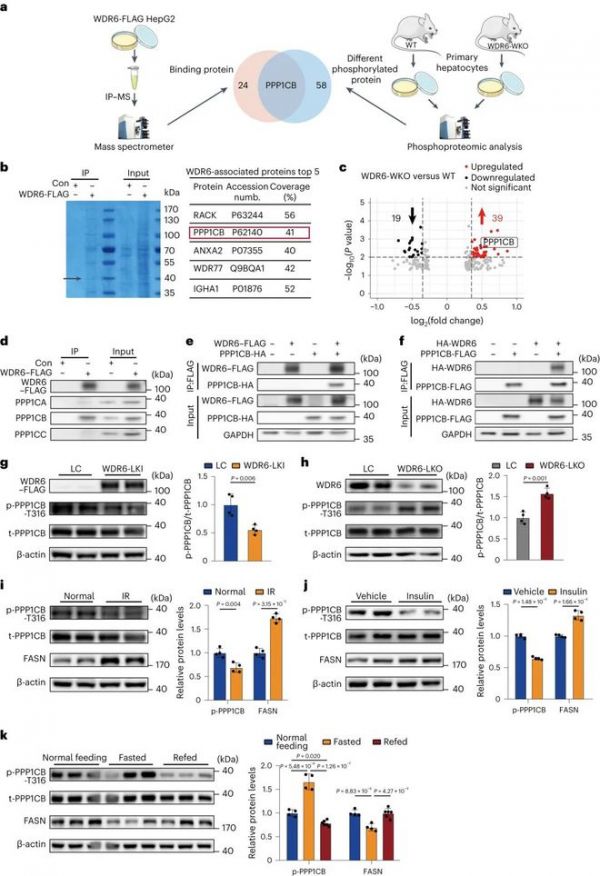
图4:WDR6与PPP1CB相互作用,促进后者在Thr316的去磷酸化
5.PPP1CB-Thr316介导WDR6调控Fasn转录
为了阐明PPP1CB在肝脏DNL中的生理功能,将si-PPP1CB或PPP1CB - flag过表达载体转染HepG2细胞。观察到PPP1CB和FASN水平之间呈负相关(图5a、b)。此外,构建了突变型PPP1CB的过表达载体,其中Thr316被丙氨酸(PPP1CB- thr316ala)或天冬氨酸(PPP1CB- thr316asp)取代,分别模拟蛋白的去磷酸化或过度磷酸化形式。与上述数据一致,我们发现过表达PPP1CB-Thr316Ala后FASN水平升高,相反,过表达PPP1CB-Thr316Asp后FASN水平降低(图5c)。然后,我们进行了体内研究,以评估PPP1CB突变体的影响。进行了aav介导的PPP1CB-Thr316Asp肝脏过表达(PPP1CB-Thr316Asp小鼠)。与对照组相比,这导致了较低的肝脏脂质沉积和TAG水平,并伴有磷酸化dna - pk增加和较低的FASN水平(图5d-f)。在此基础上,我们随后过表达WDR6,以评估当PPP1CB突变时,WDR6是否仍然可以增强DNL。与上述结果相似,WDR6表达增加加重了对照组肝脏脂质沉积,增加了FASN水平;然而,这些差异在PPP1CB-Thr316Asp小鼠中减弱(图5g-i)。此外,我们发现PP1/DNA-PK的下游靶点USF1在WDR6过表达后增加,而在PPP1CB-Thr316Asp过表达时反应减弱。这些数据表明,磷酸化ppp1cb (Thr316)介导了WDR6对USF1和FASN31的调节。由于SREBP1c不是wdr6诱导的脂肪生成调节的必要介质(图3h-j),因此我们假设USF1可能发挥这一作用。分离Usf1 - / -小鼠和WT同窝小鼠的原代肝细胞,并转染wdr6过表达腺病毒(Usf1 - / - + Ad-WDR6)或阴性对照(Usf1 - / - + Ad-GFP)。如果没有Usf1, WDR6无法驱动脂肪生成和细胞TAG沉积(图5j,k)。
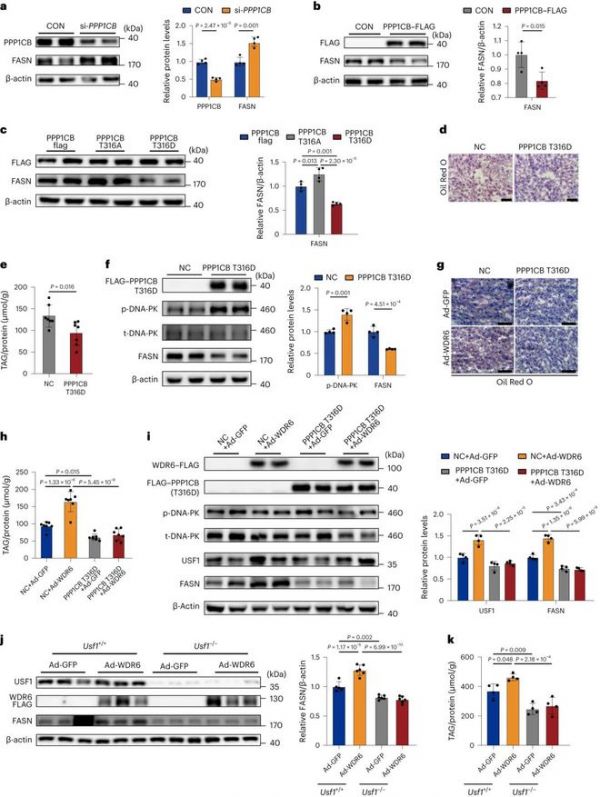
图5:WDR6通过促进PPP1CB去磷酸化调控FASN转录
6.WDR6通过FKSRSR基序与PPP1CB相互作用
为探索WDR6与PPP1CB的相互作用机制,利用AlphaFold蛋白质结构数据库进行同源建模,预测WDR6蛋白的结构。预测WDR6具有三个高度保守的β-barrel拓扑结构域(图6a)。基于预测的结构,我们设计了一系列截短的WDR6-FLAG变异体(图6b),并在HEK293细胞中分别与ha标记的PPP1CB共过表达。基于flag的IP分析显示,PPP1CB-HA在含有WDR6基因b变异型(334 ~ 687个氨基酸)和a + b变异型(1 ~ 699个氨基酸;图6c),表明WDR6通过b结构域与PPP1CB结合。值得注意的是,我们发现了一个假定的pp1结合基序“FKSRSR”(F-x-x-K/R-x- k /R,其中x可以是任何残基)位于b变异体(342-347个氨基酸;图6 d)。该基序首先在B细胞淋巴瘤2 (BCL-2)中发现,并被识别为ppp1c依赖的凋亡标记。随后,将b变异体中基序的保守氨基酸突变为丙氨酸(b-mut- flag),以确定该结构域是否是与PPP1CB结合所必需的(图6d)。IP分析表明,WDR6的b-mut-FLAG版本不能与PPP1CB结合(图6e)。
据我们所知,WD40结构域负责与其他蛋白质结合;因此,推测WDR6可能通过作为某种磷酸酶的支架蛋白来实现磷酸化ppp1cb (Thr316)的去磷酸化。迄今为止,丝氨酸/苏氨酸蛋白磷酸酶主要分布在PP1和PP2A家族。因此,我们用几种针对不同磷酸酶的抑制剂处理HepG2细胞,特别是针对PP1和PP2A家族磷酸酶。Calyculin A (PP1和PP2A41的抑制剂)以剂量依赖性方式增加了磷酸化ppp1cb (Thr316)的水平(图6f)。冈田酸也是PP1(半数抑制浓度IC50 = 10 nM)和PP2A (IC50 = 0.1 nM)的抑制剂,在1 ~ 2 nM时完全抑制PP2A活性,而抑制PP1活性需要更高的浓度42,43,44。当用5 nM或25 nM冈田酸处理细胞时,磷酸化ppp1cb (Thr316)的水平只显示出适度的增加,在这种情况下,PP2A的活性被削弱,但PP1仍然保持活性。然而,在100 nM冈田酸浓度下,磷酸化ppp1cb (Thr316)水平的增加更为明显,这是PP1活性受到抑制的浓度(图6f)。接下来,检测了转染靶向Ppp1ca, Ppp1cb或Ppp1cc sirna的小鼠原代肝细胞中磷酸化Ppp1cb (Thr316)的水平。在ppp1ca缺陷细胞中,磷酸化PPP1CB (Thr316)与总PPP1CB的比率高于PPP1CB缺陷或ppp1cc缺陷细胞(图6g-i)。这些结果表明,PPP1CA可能参与了PPP1CB (Thr316)的去磷酸化。因此,我们在WDR6-WKO小鼠和WT小鼠的原代肝细胞中对PPP1CB进行了IP。我们发现,在WT肝细胞中,PPP1CB可以与PPP1CA结合,但当WDR6缺失时,结合量减少(图6j)。综上所述,这些结果表明wdr6介导的肝脏DNL调节是通过与PPP1CB的相互作用来促进PPP1CB在Thr316的去磷酸化。
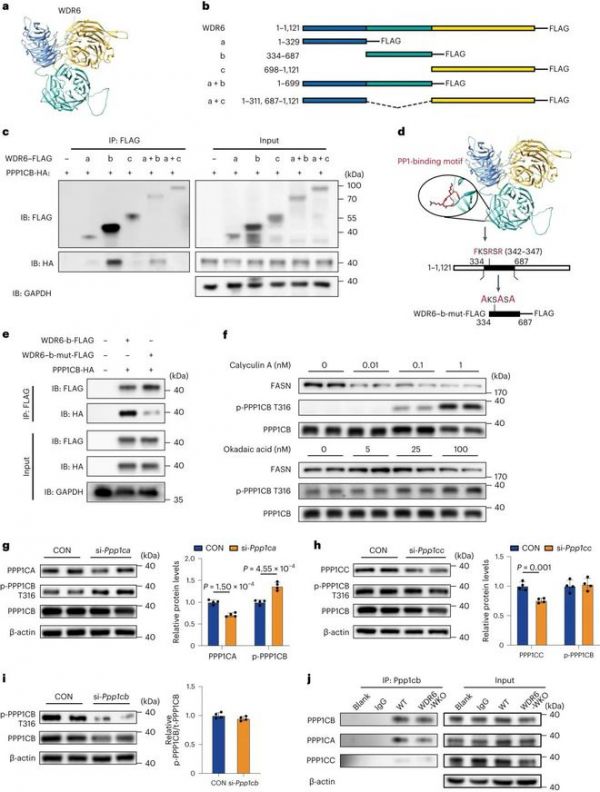
图6:WDR6的.FKSRSR基序与PPP1CB相互作用,而PPP1CA参与PPP1CB的去磷酸化
7. XLIX抑制WDR6-PPP1CB相互作用并减少肝脂肪变性
为了将我们的发现扩展到人类样本,我们检测了从2016年3月至2021年4月在山东省人民医院接受肝切除术的肝胆肿瘤患者的20个肝组织样本中WDR6和FASN的蛋白水平以及肝脏TAG含量 。组织样本(“人肝样本”)经病理学证实为非肿瘤肝组织 。在TAG水平较高的肝样本中,WDR6和FASN水平升高,而磷酸化ppp1cb (Thr316)水平较低(图7a)。肝脏WDR6蛋白水平与临床肝脏样本中的肝脏TAG或FASN水平呈正相关(图7b-d)。为了进一步研究WDR6-PPP1CB的结合模式,我们进行了计算机模拟蛋白-蛋白对接,构建了WDR6-PPP1CB复合物,并对其进行了分子动力学模拟。结果表明,WDR6的假定pp1结合基序FKSRSR(残基342-347)形成了一个可以插入到PPP1CB的一个口袋中的环(图7e)。在PPP1CB中,FKSRSR环路中的Arg345被插入到口袋中最深处(图7e)。在WDR6-XLIX复合体中,FKSRSR环与XLIX结合后构象发生明显变化。Cα原子在Arg345和Arg347中的运动距离分别为3.5 Å和8.9 Å。因此,我们推测XLIX引起的变构效应可能会干扰FKSRSR环与PPP1CB的相互作用(图7f)。
对小鼠进行了3周的HFD喂养,随后进行了4周的XLIX治疗(每天腹腔注射40 mg / kg体重)。与对照溶剂治疗组相比,xlix治疗组的血脂和胰岛素敏感性改善。此外,肝脏脂质沉积和TAG水平降低(图7g,h)。值得注意的是,磷酸化ppp1cb (Thr316)增加,FASN水平降低(图7i)。对于NASH模型,在喂食2 ~ 16周期间,在使用HFHC饮食的小鼠中,肝脏WDR6增加,同时NASH的病理进展,而磷酸化ppp1cb (Thr316)如预期的那样减少。脂质沉积和TAG水平(图7j,k)也降低。一些标志基因,如促炎相关基因Il1b、Il6和Tnf,以及促纤维化基因Col1a1、Col3a1和Acta2(编码蛋白α-SMA)的表达也降低(图7l)。这些数据表明XLIX可以减轻肝脏炎症和纤维化的进展。重要的是,与xlix处理的HFD小鼠模型相似,磷酸化ppp1cb (Thr316)的水平在xlix处理的NASH小鼠组中增加(图7m)。最后,在WDR6- lki小鼠中进行XLIX治疗,以评估XLIX是否仍然有效改善WDR6水平过高(在转录水平上约300倍过表达)小鼠的脂质沉积。我们发现,肝脏特异性Wdr6过表达减弱了XLIX减少肝脏脂质蓄积的能力(图7n,o)。综上所述,这些结果表明XLIX可以改善NAFLD和NASH的进展,可能是通过增强PPP1CB在Thr316的磷酸化,并与WDR6有关。
图7:WDR6可能是肝脂肪变性的潜在治疗靶点
结论
我们的研究支持以下观点:除了典型的胰岛素信号通路之外,还有另外一种途径可使胰岛素驱动肝脏DNL,并且WDR6表达的变化可能可以解释肝脏脂肪变性加重。然而,仍有一些局限性。首先,我们需要进一步了解机制,以详细解释高胰岛素血症如何增加WDR6表达。其次,由于PP1可以调节广泛的细胞过程,并且催化单元高度保守,因此需要测试其他PP1底物,以确定PPP1CB的Thr316是否影响PP1对这些底物的活性。最后,XLIX的具体作用机制有待阐明,XLIX对WDR6与PPP1CB结合的影响也有待进一步评估。
Yao Z, Gong Y, Chen W, Shao S, Song Y, Guo H, Li Q, Liu S, Wang X, Zhang Z, Wang Q, Xu Y, Wu Y, Wan Q, Zhao X, Xuan Q, Wang D, Lin X, Xu J, Liu J, Proud CG, Wang X, Yang R, Fu L, Niu S, Kong J, Gao L, Bo T, Zhao J. Upregulation of WDR6 drives hepatic de novo lipogenesis in insulin resistance in mice. Nat Metab. 2023 Oct;5(10):1706-1725. doi: 10.1038/s42255-023-00896-7.
注:本文编译,非声张版权,侵删!
中医药基础科研服务:
相关咨询,加微信:1278317307。
【福利时刻】科研服务(点击查看):、、、、、。咨询加微信:1278317307。

相关知识
1预防医学
1分钟读懂功能医学
社区健康与行为医学系
“康复医学”“运动医学”“运动康复”,怎么区分?
[临床医学]社区里康复.ppt
中医养生学:区域养生
《运动医学与健康科学》2023年第1期导读
随心配1+1超值登场!红区选1,白区选1,只要12元!
成都中医药大学附院针灸学校龙泉校区
生物医学的四个误区
网址: 医学1区if18+ https://www.trfsz.com/newsview856851.html
推荐资讯



- 1从出汗看健康 出汗透露你的健 3880
- 2早上怎么喝水最健康? 3664
- 3习惯造就健康 影响健康的习惯 3311
- 4五大原因危害女性健康 如何保 3219
- 5连花清瘟、布洛芬等多款感冒药 2975
- 6男子喝水喉咙里像放了刀子一样 2469
- 710人混检核酸几天出结果?1 2233
- 8第二轮新冠疫情要来了?疾控中 2232
- 9转阴多久没有传染性?满足四个 2167
- 10补肾吃什么 补肾最佳食物推荐 2166

